什么是「相分离(phase separation)」?近年来取得了哪些进展,会成为蛋白质研究的热点吗?
9 个回答

液-液相分离(phase sparation)不完全是个生物学概念,更早的时候相分离被用于物理化学中,描述两种混合后的液体相互分离的现象。
举个例子,我们往汤里滴点香油,你就能看到很多的油滴漂浮在汤表面而不是与汤互溶,形成一片一片的油花儿,这就是一种液-液相分离。


细胞内通过形成这种液-液相分离,可以形成独立的一个小环境,在其中可以独立进行生物学反应,防止无关的其他分子进入其中。
先说相变:
相变指的是物质从一种相转变为另一种相的过程,比如从固体到液体。
细胞中的相分离跟汤里的油滴不一样,它的“油滴”并不是直接由液体形成,由弱相互作用的分子(蛋白质和RNA)聚集在一起,发生相变(phase interaction),从而产生了独立的液滴。
但是生物体中的相变不会终止于液滴为止,他们可以从液体逐渐转变为固体,形成无膜细胞器。
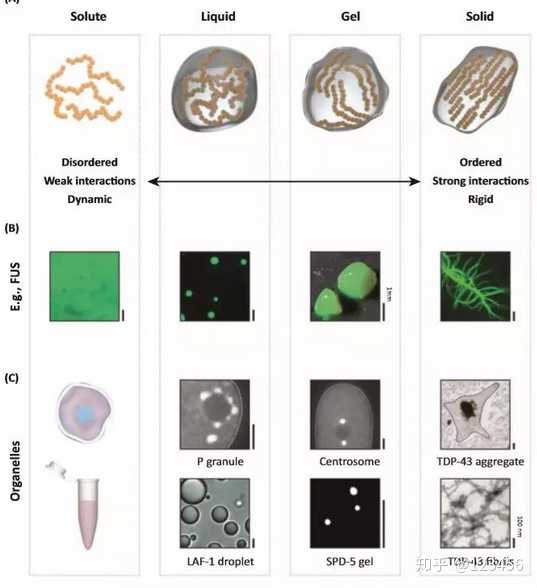
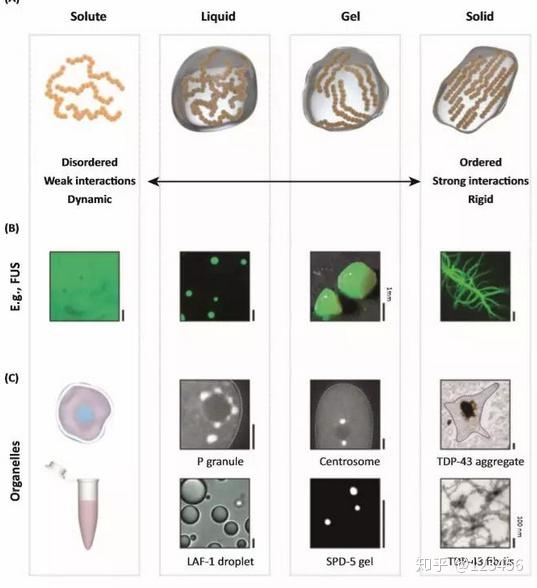
在形成这些无膜细胞器的蛋白中,广泛存在内部无序区域(intrinsically disordered regions)。这些无序区域可以作为短基序的支架,促进无膜细胞器的形成。
细胞内的相分离:
在原来的细胞概念中,细胞内环境杂乱而无序,太多的分子在其中无规律的游走,那么到底是什么帮助了生物学反应的进行,并且避免了来自那些不需要的物质的干扰呢?
通过“相分离”,也许可以提供一种方式让细胞内的特定分子聚集起来,从而在“混乱的”细胞内部形成一定“秩序”
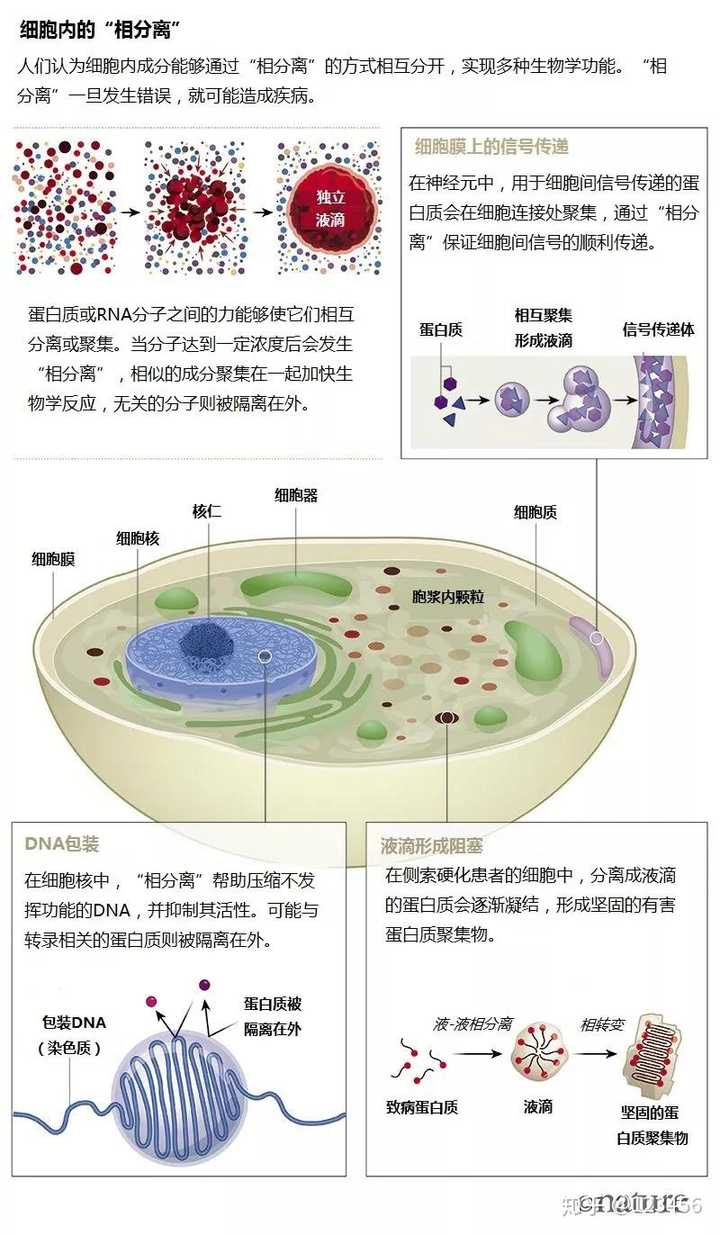
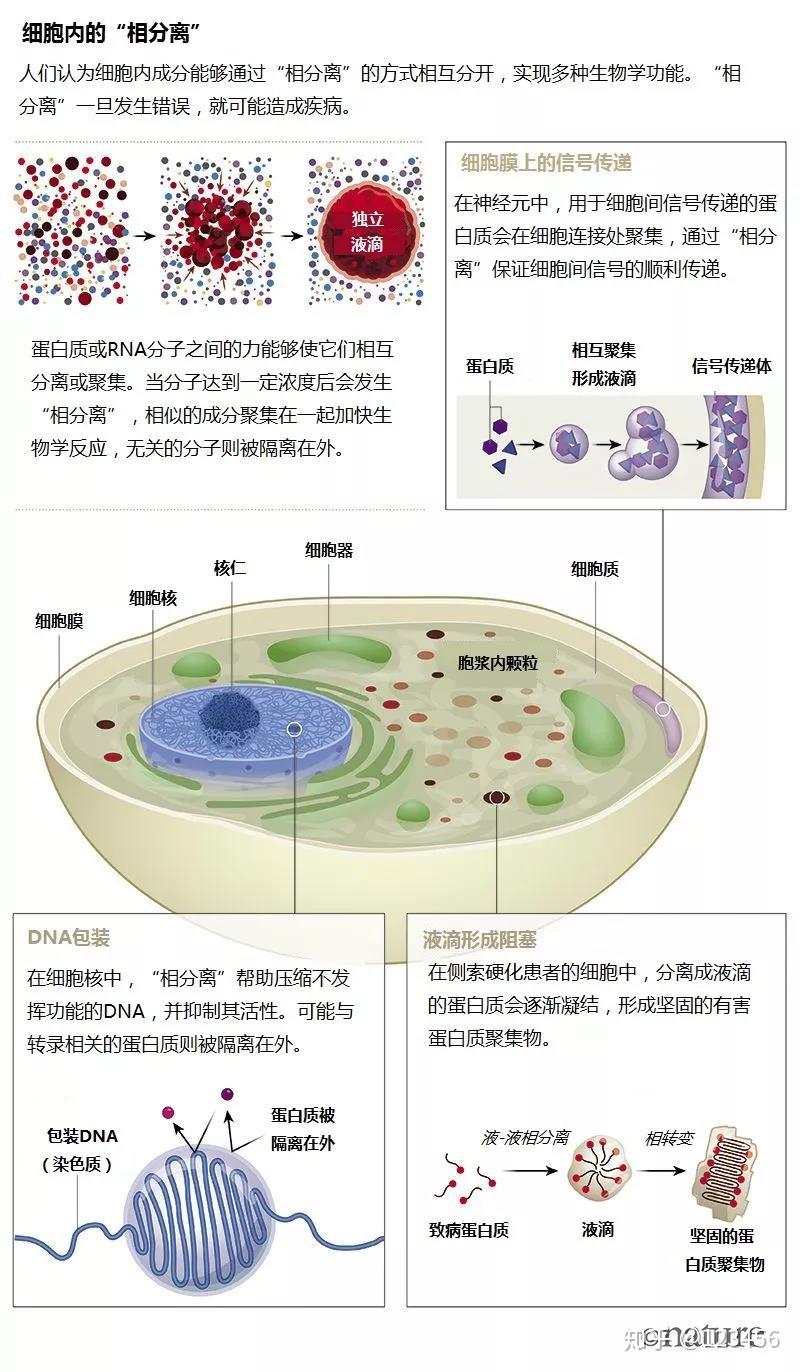
下图来自于 What lava lamps and vinaigrette can teach us about cell biology 翻译版本链接:相分离:细胞生物学的百万美元难题 这篇文章回顾了相分离的研究历史,推荐
近年来有哪些进展?
我不是做这个的,说两个我印象比较深的:
最近的最大进展应该就是相分离和转录之间的关系了,当时一出来真的是举世皆惊。
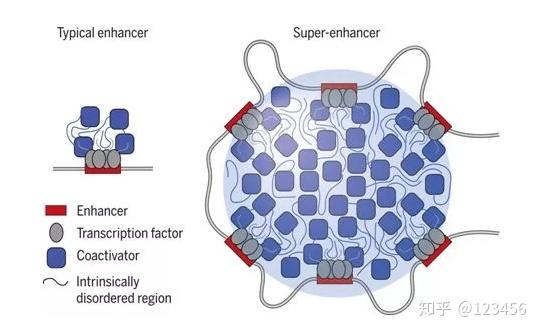
在超级增强子调控的转录复合物中,转录辅激活因子BRD4和MED1存在固有无序区域,能够形成相分离的液滴,从而调控相应基因的转录作用。

另一个是蛋白相变调控细胞命运决定因子定位,是复旦大学的温文玉老师做的。
这篇研究报道了蛋白质的相变调控了细胞命运决定因子极性聚集进而促使神经干细胞分化。
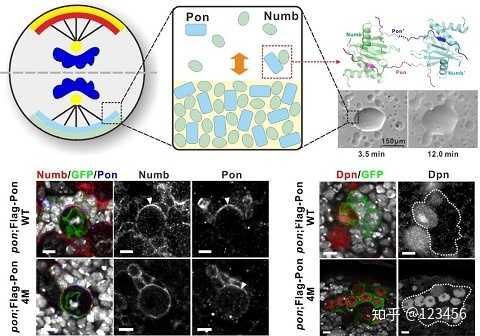
在果蝇神经干细胞(neuroblast,NB)发生不对称分裂时,命运决定因子及其衔接蛋白复合物(如Numb/Pon复合物)在底端皮层形成新月状聚集体,进而不对称分离到底部子细胞中,并促使其分化产生神经细胞。新月聚集体仅发生在分裂期,分裂结束这些蛋白又均匀分布至整个质膜或胞质中。该研究发现,Numb的PTB结构域以一种非典型的模式特异性识别Pon中的重复模序,这种多位点结合诱导该复合物发生液-液相变(liquid-liquid phase transition, LLPT)分离,在体外和细胞中能够自发组装形成致密的无膜液相结构。而这种相变液滴可以被Numb PTB的竞争多肽破坏并去组装。
顺便说一嘴:相分离/相变灌水还是挺难的,对仪器和实验设计的要求特别高,也比较难观察,门槛还是比较高的。所以虽然火,估计不会出现一窝蜂研究这个的趋势。
想写的就这些,我看这个完全是出于好玩.....有错误的话感谢各位大佬批评指正
参考:
Shan Z , Tu Y , Yang Y , et al. Basal condensation of Numb and Pon complex via phase transition during Drosophila neuroblast asymmetric division[J]. Nature Communications, 2018, 9(1):737.
Elie D . What lava lamps and vinaigrette can teach us about cell biology[J]. Nature, 2018, 555(7696):300-302.
Boeynaems S , Alberti S , Fawzi N L , et al. Protein Phase Separation: A New Phase in Cell Biology[J]. Trends in Cell Biology, 2018:S096289241830028X.

二次更新:
有关相分离的更多知识可参考今日发表于专栏:BioArt深度 上的最新文章:『珍藏版』Cell发布“相分离”研究指南 以及该文章底部的延伸阅读。
谢@李翛然 邀
本回答首发于BioArt,撰文:苏晓磊(耶鲁大学助理教授)
长文预警......


相分离(Phase Separation)的历史源远流长。早在洪荒之初,宇宙混沌一片。盘古觉得无趣,拿斧子一劈,清者上升为天,浊者下沉为地,这就是最初的相分离。作为一个经典的物理化学概念,相分离现象普遍存在于自然界中。而近期相分离又成为了生物学研究的一个热点。那么有哪些生理过程和相分离有关?相分离为什么吸引了大家的关注?怎样研究相分离? 以下内容仅作为一家之言,权当抛砖引玉。
相分离的生物学背景
在细胞水平,生物演化的一个重要特征是从相对均一的细胞质环境发展出多样化的细胞器结构。不同生物反应在不同的细胞器内发生,井井有条。比如基因转录发生在细胞核内,蛋白修饰发生在内质网和高尔基体内,分子降解发生在溶酶体内等。这些细胞器由单层或双层分子膜包围,与周围环境隔开。与此相对,细胞内有另外一类结构,称为无膜细胞器,比如核仁(nucleolus)、卡哈尔体(Cajal body)和应激颗粒(stress granule)等。这些结构稳定存在,但是没有细胞膜包裹,仍与周围环境产生频繁的分子交换。它们为什么能稳定存在成为了一个有趣的问题。而相分离提供了一个形成此类细胞器的机制。某些蛋白质或者核酸分子可以通过多价相互作用,在原本均一的环境中产生物理化学性质不同的另一相,形成无膜细胞器或者是细胞结构。在绝大多数情况下,这些细胞结构呈现液态特征,所以被称为液滴 (liquid droplet) 或者是液态凝聚体 (liquid condensates)。
相分离形成的细胞结构有哪些?功能是什么?
相分离形成的结构大致可分为二维和三维的。二维结构主要存在于细胞膜上,比如肾过滤屏障组成蛋白nephrin聚集体,能诱导微丝聚合 【1,2】 ; T细胞微体 (T cell microcluster),促进T细胞活化【3】 ;神经细胞后突触质密区(PSD),稳定神经突触结构【4,5】。三维相分离结构存在于细胞质和细胞核内。比如细胞质内的P颗粒,参与线虫早期胚胎发育 【6】;应激颗粒(stress granule), 在细胞受到胁迫条件下产生,有利于胁迫解除后细胞快速恢复常态【7,8】;再比如细胞核内促进rRNA加工的核仁【9】,某些转录调控的超级增强子【10】,异染色质【11,12】也被报道形成相分离结构; 同时,RNA干涉【13】,纺锤体形成【14】,神经细胞不对称分裂【15】,细胞自噬【16,17】,先天免疫应答 (Du M, Science 2018)【18】,光合作用 【19】等多种过程中都有相分离结构形成。近几年的研究表明,相分离成为了一种较为普遍形成细胞结构的机制,在细胞各项生理过程中广泛存在。
相分离为什么引起广泛关注?
相分离为研究生物分子“高维”结构和功能提供了一个新视点。单个蛋白质分子的三维结构决定其物理化学特性;蛋白复合体的形成,可以实现很多生物学功能。而相分离描述了更高一个层次的结构,尺寸在微米级别。它关注的不仅仅是几个关键蛋白本身的特性,更着重于创造一个相对独立的空间域,选择性地富集分子并组成一定结构。通常相分离形成的细胞结构与周围环境相比,有更高的蛋白密度以及减弱的分子微观运动,这可以促进某些生化反应。同时,相分离结构也可以通过限制分子运动,隔离底物,抑制反应发生。因时因地而异,相分离结构的作用都会有所不同。
相分离也为解释某些细胞骤变现象 (switch-like behavior) 提供了分子依据。相分离可以通过相变(phase transition) 发生。相变的过程一般很急速。如果处于临界态附近,一个微小的环境变化能够引起从0到1的巨变。比如说,T细胞受体如何分辨自身抗原和外界抗原(细菌、病毒、肿瘤等)是一个研究了几十年的课题,至今没有明确答案。自身抗原和外界抗原与T细胞受体的亲和性相差不大,为什么自身抗原不能激活T细胞而外界抗原可以?而在T细胞内最近发现的相分离过程,则有望为解释T细胞活化过程中的骤变提供机理。由于T细胞在肿瘤免疫疗法中的重要作用,相关研究也可能为发展新的T细胞疗法提供靶点。
我研究的蛋白会不会相分离?
正如每个人都有爱的权利,每一个蛋白都有相分离的可能。事实上,结构生物学中蛋白结晶也是一个相分离过程,只不过这样的相分离一般发生在非生理条件下。所以问题的关键不是能否相分离,而是在生理相关的条件下,是否会发生相分离。
同时,生理相关的条件起变化时,也可以改变相分离形成的稳态。在细胞内,这可以是饥饿引起的pH值降低,粘度变化,也可以是在细胞激活情况下钙离子或其他高价阳离子浓度上升,还可以是蛋白本身的表达水平变化,磷酸化修饰等等。对于非恒温生物,如酵母、线虫等,温度的改变也可能引发胞内蛋白相分离。观察生理相关条件变化前后蛋白聚集状态的变化,这也许是发现相分离的关键一步。
如何鉴定相分离?
用GFP标记一个蛋白,发现在细胞中形成球状的小点,这是不是就说明是相分离了呢?要是这样想的话未免就有些 too young too simple 了。综合几篇综述(Hyman AA Annu Rev Cell Dev Biol. 2014; Banani SF Nat Rev Mol Cell Biol. 2017; Berry J Rep Prog Phys. 2018)【20-22】,要确定是不是所研究的是不是液态相分离结构(细胞中最为常见), 需要几方面的实验证据:1.FRAP(光脱色荧光恢复技术,fluorescence recovery after photobleaching ),看所研究的蛋白是否在短时间内恢复荧光。这可以表明此种结构是否与周围环境在进行频繁的物质交换;2. 结构大小是否达到或接近微米级别;3.活细胞成像,是否能观察到液滴融合或分离现象;4. 体外重构。利用纯化的蛋白,在生理相关的条件下是否可以重构出相分离结构。这也是确定相分离机制的有效手段。
相分离领域的未来关键问题
- 相分离的精细结构
由于液态特性,细胞中的相分离结构往往没有稳定的组成和构象,很难用主流的结构生物学方法如晶体学或者是单颗粒电镜来研究。超分辨显微术可以解析亚结构和分子组成,但是分辨率只能接近分子水平,无法深入到蛋白二级结构。因而相分离的精细结构需要靠发展新的技术来解析。
2. 相分离的生物学功能
目前对于相分离所形成的细胞器的结构和机理已经有了许多认识,接下来的主要目标是研究相分离的生物学功能。在生化层面,相分离结构可以选择性地富集或者是排斥分子,增加分子相互作用几率,甚至改变分子构象以促进或者是抑制化学反应。将这些物理和化学性质与生物学功能相匹配,将是今后研究的一个重要问题。另外,以相分离为靶点,改造细胞生理过程,比如信号转导、转录调控、应激反应等等,也将为相分离研究提供广阔的应用前景。
3. 相分离与疾病的关系
这一方面的研究方兴未艾。目前的研究主要集中在神经退行性疾病。某些蛋白点突变,如FUS能使通常液态的相分离结构加速变为固态,这可能是神经退行性疾病中常见的不容聚集体形成的原因 (Wang J, Cell 2018)【23】;此外,入核转运可以降低细胞质内核酸结合蛋白浓度,溶解液态相分离结构,防止固态凝聚体造成的神经细胞毒性 (Guo L, Cell 2018;Hofweber M,Cell 2018;Yoshizawa T,Cell 2018)【24-26】。另外,基因组短小重复序列增生也和神经退行性疾病相关。有研究表明,这些重复序列转录成的RNA也能够形成相分结构,这可能是致病的原因之一 (Jain A, Nature 2017)【27】。预计今后相分离相关的疾病研究将从神经退行性疾病的研究扩展到癌症,免疫等方面。有关相分离机制的研究,特别是寻找或者是筛选调节相分离的关键分子,如激酶,将为确定有效靶点分子提供理论依据。
注:感谢清华大学李丕龙教授阅读本文并提出修改意见。
作者简介
苏晓磊,哈佛大学细胞与发育生物学博士。现耶鲁大学细胞生物学助理教授。研究方向为免疫细胞中的膜生物学,尤其是相分离在T细胞活化中的作用。欢迎对相分离和免疫学感兴趣的同学加入团队。(https://www.sulab.net/recruitment)
参考文献:
1、Li, P., Banjade, S., Cheng, H. C., Kim, S., Chen, B., Guo, L., ... & Russo, P. S. (2012). Phase transitions in the assembly of multivalent signalling proteins. Nature, 483(7389), 336.
2、Banjade, S., & Rosen, M. K. (2014). Phase transitions of multivalent proteins can promote clustering of membrane receptors. Elife, 3, e04123.
3、Su, X., Ditlev, J. A., Hui, E., Xing, W., Banjade, S., Okrut, J., ... & Vale, R. D. (2016). Phase separation of signaling molecules promotes T cell receptor signal transduction. Science, 352(6285), 595-599.
4、Zeng, M., Shang, Y., Araki, Y., Guo, T., Huganir, R. L., & Zhang, M. (2016). Phase transition in postsynaptic densities underlies formation of synaptic complexes and synaptic plasticity. Cell, 166(5), 1163-1175.
5、Zeng, M., Chen, X., Guan, D., Xu, J., Wu, H., Tong, P., & Zhang, M. (2018). Reconstituted Postsynaptic Density as a Molecular Platform for Understanding Synapse Formation and Plasticity. Cell.
6、Brangwynne, C. P., Eckmann, C. R., Courson, D. S., Rybarska, A., Hoege, C., Gharakhani, J., ... & Hyman, A. A. (2009). Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science, 324(5935), 1729-1732.
7、Molliex, A., Temirov, J., Lee, J., Coughlin, M., Kanagaraj, A. P., Kim, H. J., ... & Taylor, J. P. (2015). Phase separation by low complexity domains promotes stress granule assembly and drives pathological fibrillization. Cell, 163(1), 123-133.
8、Riback, J. A., Katanski, C. D., Kear-Scott, J. L., Pilipenko, E. V., Rojek, A. E., Sosnick, T. R., & Drummond, D. A. (2017). Stress-triggered phase separation is an adaptive, evolutionarily tuned response. Cell, 168(6), 1028-1040.
9、Feric, M., Vaidya, N., Harmon, T. S., Mitrea, D. M., Zhu, L., Richardson, T. M., ... & Brangwynne, C. P. (2016). Coexisting liquid phases underlie nucleolar subcompartments.Cell, 165(7), 1686-1697.
10、Sabari, B. R., Dall'Agnese, A., Boija, A., Klein, I. A., Coffey, E. L., Shrinivas, K., ... & Li, C. H. (2018). Coactivator condensation at super-enhancers links phase separation and gene control. Science, eaar3958.
11、Strom, A. R., Emelyanov, A. V., Mir, M., Fyodorov, D. V., Darzacq, X., & Karpen, G. H. (2017). Phase separation drives heterochromatin domain formation. Nature, 547(7662), 241.
12、Larson, A. G., Elnatan, D., Keenen, M. M., Trnka, M. J., Johnston, J. B., Burlingame, A. L., ... & Narlikar, G. J. (2017). Liquid droplet formation by HP1α suggests a role for phase separation in heterochromatin. Nature, 547(7662), 236.
13、Sheu-Gruttadauria, J., & MacRae, I. J. (2018). Phase Transitions in the Assembly and Function of Human miRISC. Cell, 173(4), 946-957.
14、Jiang, H., Wang, S., Huang, Y., He, X., Cui, H., Zhu, X., & Zheng, Y. (2015). Phase transition of spindle-associated protein regulate spindle apparatus assembly. Cell, 163(1), 108-122.
15、Shan, Z., Tu, Y., Yang, Y., Liu, Z., Zeng, M., Xu, H., ... & Wen, W. (2018). Basal condensation of Numb and Pon complex via phase transition during Drosophila neuroblast asymmetric division. Nature communications, 9(1), 737.
16、Sun, D., Wu, R., Zheng, J., Li, P., & Yu, L. (2018). Polyubiquitin chain-induced p62 phase separation drives autophagic cargo segregation. Cell research, 1.
17、Zhang G, mTOR regulates phase separation of PGL granules to modulate their autophagic degradation.Cell 2018
18、Du, M., & Chen, Z. J. (2018). DNA-induced liquid phase condensation of cGAS activates innate immune signaling. Science, eaat1022.
19、Rosenzweig, E. S. F., Xu, B., Cuellar, L. K., Martinez-Sanchez, A., Schaffer, M., Strauss, M., ... & Wingreen, N. S. (2017). The eukaryotic CO 2-concentrating organelle is liquid-like and exhibits dynamic reorganization. Cell, 171(1), 148-162.
20、Hyman, A. A., Weber, C. A., & Jülicher, F. (2014). Liquid-liquid phase separation in biology. Annual review of cell and developmental biology, 30, 39-58.
21、Banani, S. F., Lee, H. O., Hyman, A. A., & Rosen, M. K. (2017). Biomolecular condensates: organizers of cellular biochemistry. Nature reviews Molecular cell biology, 18(5), 285.
22、Berry, J., Brangwynne, C. P., & Haataja, M. (2018). Physical principles of intracellular organization via active and passive phase transitions. Reports on Progress in Physics, 81(4), 046601.
23、Wang, J., Choi, J. M., Holehouse, A. S., Lee, H. O., Zhang, X., Jahnel, M., ... & Poser, I. (2018). A molecular grammar governing the driving forces for phase separation of prion-like RNA binding proteins. Cell, 174(3), 688-699.
24、Guo, L., Kim, H. J., Wang, H., Monaghan, J., Freyermuth, F., Sung, J. C., ... & Zhang, Z. C. (2018). Nuclear-import receptors reverse aberrant phase transitions of RNA-binding proteins with prion-like domains. Cell, 173(3), 677-692.
25、Hofweber, M., Hutten, S., Bourgeois, B., Spreitzer, E., Niedner-Boblenz, A., Schifferer, M., Ruepp, M., Simons, M., Niessing, D., Madl, T., and Dormann, D. (2018). Phase separation of FUS is suppressed by its nuclear import recep- tor and arginine methylation. Cell 173.
26、Yoshizawa, T., Ali, R., Jiou, J., Fung, H.Y.J., Burke, K.A., Kim, S.J., Lin, Y., Peeples, W.B., Saltzberg, D., Soniat, M., et al. (2018). Nuclear import receptor inhibits phase separation of FUS through binding to multiple sites. Cell 173.
27、Jain, A., & Vale, R. D. (2017). RNA phase transitions in repeat expansion disorders. Nature, 546(7657), 243.