
Definición de replicación viral
- La replicación viral es el proceso de formación de virus biológicos en el curso del proceso de infección dentro de las células huésped.
- Un virus tiene que pasar por el proceso de reproducción para generar nuevos viriones infecciosos que puedan infectar las células dentro del cuerpo y los huéspedes posteriores.
- Después de ingresar al cuerpo, el virus hace contacto físico y atraviesa su membrana plasmática con la célula objetivo.
- En su interior crea y reproduce su genoma y facilita la producción de sus proteínas a través de los ribosomas del huésped.
- Las partículas de un virus se derivan de las biomoléculas que han sido sintetizadas y se transforman en virus infecciosos.
- Luego, los viriones son expulsados de las células para continuar con el proceso de infección.
Papel del ácido nucleico - transfección
Las células de los organismos superiores pueden verse afectadas por el ácido nucleico viral que produce iones virales normales. Hay muchas distinciones importantes entre la infección causada por el ácido nucleico (transfección) y los virus.
- La efectividad de la infección por ácido nucleico es menor, con una relación de 10-6-10-8 en medios convencionales, lo que demuestra la importancia de la cubierta de virus en la infectividad. La eficacia del virus se puede mejorar mediante la dispersión del ADN viral en las células mediante la inyección de fosfato de calcio o el empaquetamiento en liposomas.
- El rango de huéspedes es más amplio con ácidos nucleicos que son capaces de infectar las células que son resistentes, por ejemplo, las células de pollo, a pesar de no ser susceptibles al poliovirus ya que no tienen receptores para el virus, son vulnerables a su ARN pero solo a un ciclo de multiplicación por poliovirus. Ocurre porque los progenitores también son viriones y no pueden transmitirse a otras células debido a la falta de receptores apropiados u otros factores.
- Los ácidos nucleicos infecciosos se pueden extraer de virus que han sido activados por calor donde se desnaturalizan las proteínas de la cápside. Los ácidos nucleicos pueden soportar temperaturas más extremas que las proteínas. La capacidad de infectividad de los ácidos nucleicos para resistir cualquier daño causado por la cubierta del virus debe tenerse en cuenta al preparar vacunas contra virus.
- En el caso de algunos virus de ARN, las copias de ADN del ARN del virus son infecciosas. por ejemplo, poliovirus, que permite la producción de genomas virales como cepas de vacunas en grandes cantidades porque evita la alta tasa de mutación en la replicación del ARN, y también su capacidad de replicación.
- La capacidad de los ácidos nucleicos para infectar no se ve afectada por virus que son AB específicos. Esto sugiere que este tipo de virus puede ser un poderoso agente infeccioso en presencia del sistema inmunitario. Pero, las nucleasas presentes en el cuerpo podrían restringir el papel de las nucleasas. De los virus animales, los adenovirus y papovavirus así como los herpesvirus así como los togavirus y picorna generan ácido nucleico infeccioso. Los retrovirus pueden causar ADN infeccioso. puede aislarse de células infectadas o crearse copiando el ADN viral en el laboratorio.
Pasos de replicación viral
Todos los virus deben pasar por los siete pasos para producir nuevos viriones. Ciertas etapas pueden ocurrir junto con otras etapas o incluso realizarse fuera de secuencia según el virus. 7 etapas para la replicación están en el siguiente orden:
- Adjuntar archivo
- Penetración
- Destapando
- Replicación
- Asamblea
- Maduración
- tortugitas
*Nota: Una buena regla mnemotécnica para recordar las etapas de la replicación viral es la frase “UN PURple Apple Mvuelo Redén.” Las letras en negrita son los caracteres iniciales de cada una de las etapas, en orden.
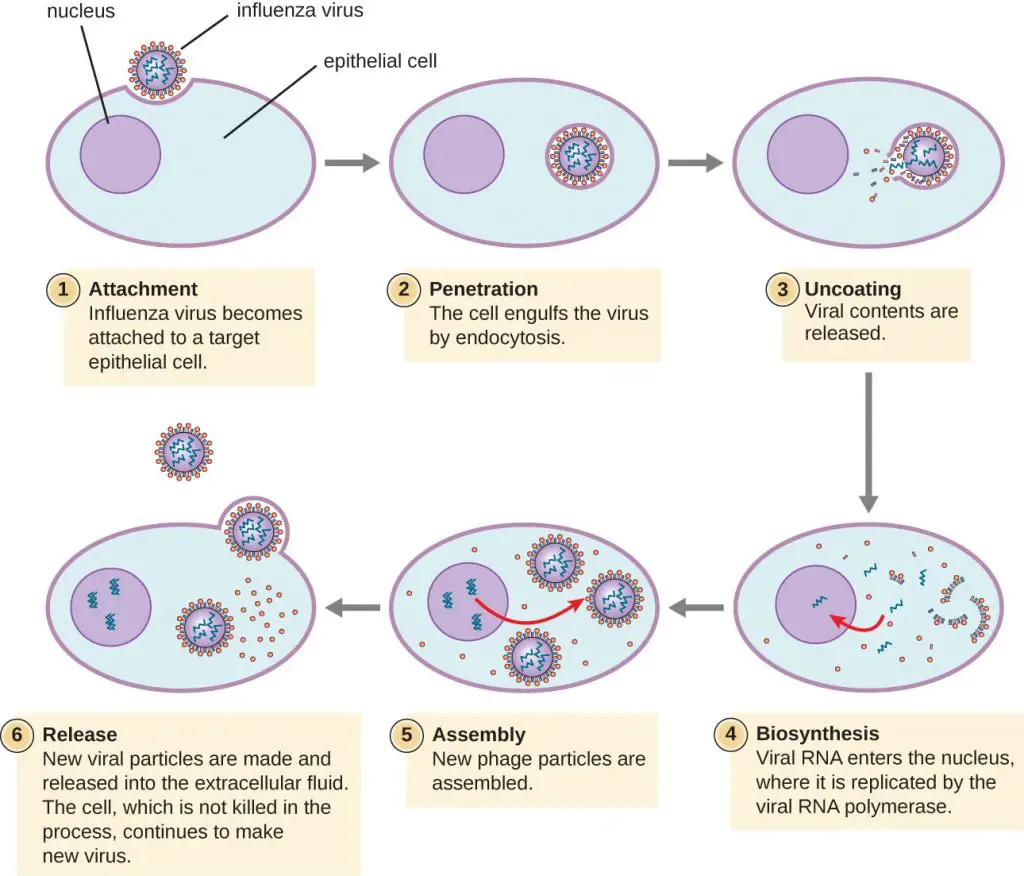
1. Fijación de partículas virales
- Es el paso inicial en el proceso de replicación viral que el virus se adhiere a las membranas de las células en la célula del huésped.
- Las membranas plasmáticas de las células se componen de una bicapa de fosfolípidos que se caracteriza por una serie de proteínas que sobresalen a través de la membrana. Las proteínas de superficie realizan diversas funciones. Estos incluyen el transporte de moléculas e iones, además de facilitar el proceso de unión de una célula a otra y actuar como receptores de las proteínas entrantes. La mayoría de las proteínas de la membrana plasmática pueden describirse como glicosiladas, lo que significa que han sido modificadas por carbohidratos y azúcares.
- Las moléculas receptoras que se dirigen a la superficie de la célula son proteínas típicas requeridas para las funciones celulares que los virus han desarrollado para aprovechar, típicamente glicoproteínas o residuos de azúcar y carbohidratos que se encuentran en las glicoproteínas o en la membrana plasmática.
- Ciertos virus también requieren correceptores para poder infectar células. Inicialmente, el VIH se adhiere a una proteína denominada CD4 en la superficie de los linfocitos T ("células T"); sin embargo, requiere una de dos proteínas conocidas como correceptores para mantener el proceso de infección.
Mecanismo de unión de células virales
- La unión resulta de fuerzas electrostáticas opuestas a la proteína de unión del virus y los receptores de superficie de las células. La proteína de unión para el virus se puede encontrar en la parte exterior del virus porque es allí donde se produce el contacto con la célula.
- La proteína de unión sobresale en la envoltura de un virus con envoltura, a diferencia de los virus sin envoltura, que tienen al menos una proteína de la cápside que se comunica con los receptores de la superficie celular.
- Las proteínas de unión del virus pueden extenderse desde el exterior del virión, o pueden estar contenidas dentro de “cañones” formados por proteínas de la cápside.
Receptores de superficie celular
Diferentes virus utilizan receptores específicos en la superficie celular para adherirse.
- Para tratar el VIH-1 es necesario que CD4 funcione como receptor y los receptores de quimiocinas CCR5 o CXCR4 como correceptores.
- El virus de la influenza se une a los residuos de ácido siálico en la terminal ubicada en las glicoproteínas de la superficie celular.
- Las cepas de laboratorio del virus del sarampión pueden unirse al CD46 (aunque el CD150 también actúa como receptor del virus del sarampión).
- El virus del herpes simple 1 primero se une al azufre de heparán en los GAG para conectarse a los receptores de entrada, por ejemplo, HVEM y nectinas.
- El noventa por ciento de los rinovirus emplean ICAM-1 como receptor, mientras que el 10% de los rinovirus utilizan ese receptor VLDL.
- Dengue virus is transmitted through DC-SIGN.
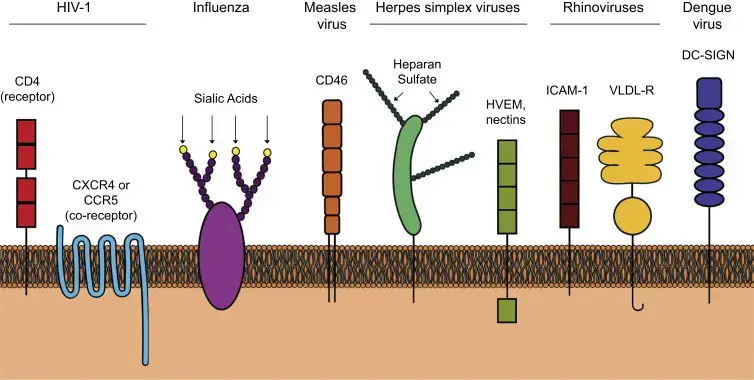
Celular Surface Receptors for Attachment of Human Viruses
| Virus | Celular surface receptor(s) |
| Rinovirus | Molécula de adhesión intercelular 1 (ICAM-1) (90 %), receptor de lipoproteínas de baja densidad (10 %) |
| Poliovirus | Receptor de poliovirus (PVR) CD155 |
| Virus de inmunodeficiencia humana | CD4 (receptor); CCR5 o CXCR4 (correceptores) |
| Virus de la influenza A | Ácido siálico |
| Virus del sarampión | CD46, CD150 |
| Virus del herpes simple-1 | Heparán sulfato, HVEM, Nectin-1 |
| Dengue virus | SIGNO DC |
| Virus de la hepatitis B | Polipéptido cotransportador de taurocolato de sodio |
| Virus del papiloma humano | Heparán sulfato, integrinas |
2. Penetración
- Después de la unión, el virus exitoso ingresa rápidamente a la célula, lo que le permite escapar de cualquier estrés extracelular que pueda dañar el virión, por ejemplo, el movimiento de la mucosidad.
- La penetración es el proceso de atravesar la membrana plasmática causado por virus. Contrariamente al apego al virus, la penetración requiere energía, pero esta es proporcionada a través de las células del huésped, pero no por el virus.
Mecanismo de Penetración Viral
Diferentes virus utilizan diversos mecanismos celulares para ingresar a la célula uniéndose a receptores específicos en la superficie celular.
- Endocitosis mediada por receptor: Receptor-mediated Endocyto is typically utilized by viruses to enter the membrane of plasma. When the pH of an endosome decreases, virus proteins alter their configuration that allows them to be released out of the endosome. Based on what virus is at play, it could occur in early endosomes, the late endosomes or lisosomas. Both enveloped and non-enveloped viruses benefit from receptor-mediated endocytosis for access into the cytoplasm the cell. Both enveloped and non-enveloped viruses make use of receptor-mediated endocytosis within pits that are coated with caveolin or clathrin.
- Endocitosis mediada por clatrina: La mayoría de los virus utilizan endocitosis mediada por clatrina para entrar en células como el virus del dengue, el virus de la hepatitis C y los reovirus.
- Endocitosis mediada por caveolas: Un puñado de virus conocidos que afectan a los seres humanos, como el SV40 y los virus del papiloma (que causan verrugas y cáncer de cuello uterino) utilizan endocitosis mediada por caveolas y esto se descubrió utilizando un medicamento que bloqueaba el desarrollo de caveolas. La eliminación de la endocitosis mediada por clatrina no detuvo la entrada del virus en la célula. Algunos virus también están sujetos a endocitosis mediada por receptores que son indistinguibles tanto de la caveolina como de la clatrina.
- Endocitosis en fase masiva: Durante la endocitosis de fase masiva, la célula crea una vesícula envolvente para engullir cualquier molécula que se encuentre en el líquido extracelular, que incluye virus.
- Fagocitosis: Fagocitosis is a form of endocytosis that is mediated by receptors and is used by cells that are specialized to completely engulf cells. Recently, two massive DNA-based viruses, including VHS-1 as well as Mimivirus were found to enter cells via phagocytosis pathways.
- Fusión: La fusión es un método de penetración que solo utilizan los virus envueltos, es la fusión. La fusión de la envoltura viral es posible en la membrana celular o en vesículas endocitadas como el endosoma. Está controlado por la misma proteína vírica utilizada por el virus para adherirse y adherirse, o mediante una proteína vírica distinta, basada en el virus.
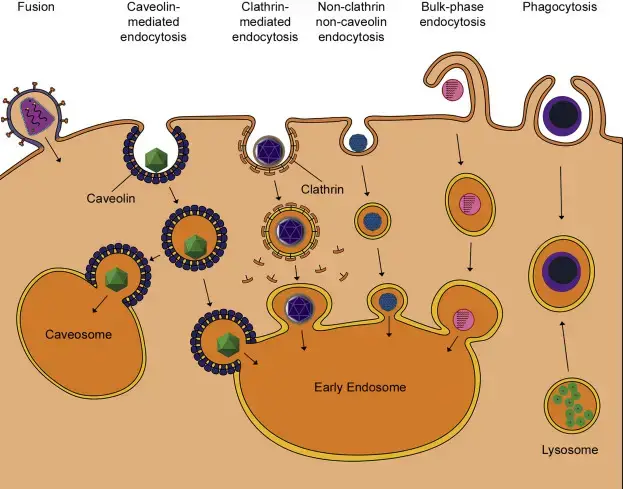
| Tipo de penetración (entrada) | Virus ejemplos |
| Endocitosis mediada por clatrina | Dengue virus, hepatitis C virus, reovirus, adenovirus, parvovirus B19, West Nile virus |
| Endocitosis mediada por caveolina | Virus del papiloma humano, SV40, virus de la hepatitis B |
| Fusion | VIH, influenza, virus respiratorio sincitial, virus del herpes simple, virus del dengue, virus del Ébola |
3. Decapado
- Uncoating es el término utilizado para describir la descomposición o eliminación de la cápside que provoca un éxodo del genoma del virus a la célula, el lugar donde se produce la replicación y la transcripción del genoma.
Mecanismo de decapado
El decapado es un proceso que puede distinguirse o estar estrechamente relacionado con la penetración. Los virus también pueden lograr el proceso de decapado a través de una variedad de métodos diferentes.
- Rinovirus: Los rinovirus se transportan a las células a través de endocitosis mediada por receptores que son clatrina-co. Dentro de los endosomas ácidos, el virus crece en tamaño en un cuatro por ciento. Una proteína de la cápside, la VP1 (proteína viral 1) es capaz de formar poros dentro del endosoma, lo que permite la liberación del genoma del rinovirus.
- Virus de la gripe: El virus de la influenza contiene una proteína específica del virus conocida como hemaglutinina (HA) en la cubierta del virus. Se une a residuos de ácido siálico en la superficie exterior de la célula epitelial respiratoria y entra en la célula a través de endocitosis mediada por receptores. La caída del pH del endosoma da como resultado cambios en la forma de la proteína HA del virus, que revela un péptido de fusión que acerca las dos membranas y conecta la envoltura viral con la membrana del endosoma. En este caso, la proteína HA ayuda en la fijación y eliminación del virus. Los segmentos del genoma del ADN viral se transportan al núcleo antes de entrar a través de los poros nucleares.
- Otras cápsides virales (Poliovirus): Se cree que la cápside del poliovirus no puede ingresar a las células de ninguna manera: la presencia del poliovirus de la cápside con el receptor de la superficie celular da como resultado una alteración en el virión, lo que crea una abertura dentro de la membrana celular a través de la cual el ARN viral puede liberarse en el citoplasma de las células. Muchos virus mantienen una cápside intacta dentro del citosol que sirve como "base de operaciones" para la replicación similar a los reovirus.
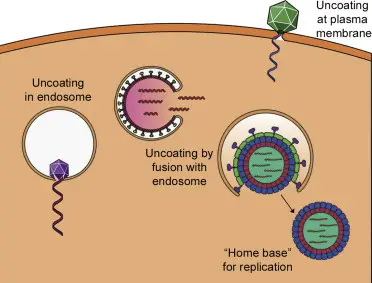
Transporte de genomas virales al núcleo.
Una variedad de virus necesitan transportar sus genomas al núcleo para que se produzca la replicación o la transcripción. Los segmentos del genoma de la gripe se transportan a través del poro nuclear hasta el núcleo. Las cápsidas del virus del herpes se transportan a través de microtúbulos al poro nuclear, desde donde se produce el desprendimiento. Las cápsides del adenovirus se rompen en el poro nuclear y el ADN del virus se transporta al núcleo. Otros virus, como el virus de la hepatitis B, son tan pequeños que toda la cápside puede atravesar los poros nucleares.
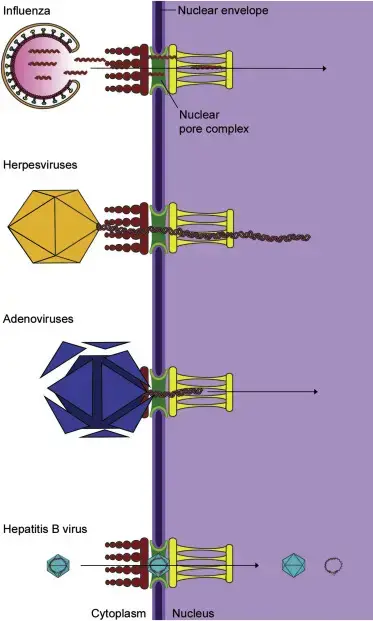
4. Replicación
Los virus virales, ya sean ADN o ARN, interrumpen el proceso de síntesis de proteínas en las células y descomponen los polirribosomas celulares, lo que conduce a un cambio hacia la síntesis de proteínas virales. El mecanismo para detener la síntesis de proteínas varía según la familia de virus.
El poliovirus, utilizando una enzima que es viral, desencadena la escisión de una proteína de unión a caperuza de 200 Kd que es esencial para el inicio del proceso de traducción de mensajeros celulares caperuza. A diferencia de los virus con virulencia, los virus leves, por ejemplo, los poliomavirus pueden estimular la producción de proteínas y ADN y ARNm del huésped. Este fenómeno es de particular interés para la carcinogénesis viral.
Virus de ADN
Para los virus de ADN en animales, la transcripción y la traducción no se conectan. A excepción de los poxvirus, la transcripción tiene lugar dentro del núcleo y la traducción dentro de las células. En general, las primeras transcripciones producidas a partir de la ARN polimerasa II son mayores que los ARNm de los ribosomas. En algunos casos, existe la posibilidad de que hasta el 70 % del ARN transcripcional no se traduzca en el núcleo.
Los mensajeros del virus pero, como ocurre con las células animales, pueden ser unicéntricos. La transcripción tiene una estructura temporal ordenada, y para la mayoría de los virus de ADN, solo una pequeña fracción de su genoma se transcribe en los primeros mensajes. La síntesis de proteínas en las primeras etapas es la etapa primaria en la replicación del ADN viral.
Después de la síntesis de ADN, el genoma restante se transcribe en mensajes de etapa tardía. Los virus complejos poseen genes tempranos inmediatos que se activan en presencia de proteínas que inhiben la producción de proteínas, así como genes tempranos retrasados que requieren la expresión de la síntesis de proteínas.
La regulación se logra a través de proteínas que se encuentran en el virus, o según lo especificado por genes celulares o virales, que interactúan con secuencias reguladoras ubicadas en el extremo 5' del gen. Estas secuencias podrían reaccionar en trans a los productos generados por otros genes y también actuar en cis con los genes asociados a ella. Se pueden transcribir diferentes tipos de genes a partir de diferentes hebras de ADN y, por lo tanto, en orientaciones opuestas, por ejemplo, poliomavirus. Las transcripciones pueden someterse a un procesamiento postranscripcional para garantizar que se eliminen las secuencias intermedias no esenciales.
Replicación de ADN
El método de replicación puede describirse como semiconservador, sin embargo, los intermediarios que se replican dependen del modo en que se lleva a cabo la replicación. Se pueden identificar varios métodos de replicación.
- Adenovirus: Los adenovirus tienen una replicación asimétrica que comienza en el extremo 3' de una de las cadenas, utilizando el cebador proteico. La cadena creciente desplazará a la anterior con la misma polaridad y creará la estructura doble completa. La hebra desplazada luego se replica de manera similar al formar un mango al combinarse con las repeticiones terminales invertidas.
- Herpesvirus: Tiene genomas lineales que tienen repeticiones terminales. Al llegar al núcleo, estos terminales sufren una digestión exonucleótica limitada y luego se forman circulares. Se cree que la replicación ocurre a través del mecanismo de círculo rodante en el que se crean los concatemeros. Durante la maduración, las moléculas de longitud unitaria se separan de los concatemeros.
- Papovavirus: La secuencia de ADN de los papovavirus es circular y el proceso de replicación es bidireccional y simétrico a través de intercalaciones que son cíclicas.
- Parvovirus: La replicación de los parvovirus monocatenarios comenzó cuando el ADN de cadena +ve y -ve de varios parvovirus se unieron para formar una molécula de ADN de doble cadena. Es a partir de esto que se lleva a cabo la replicación y la transcripción.
- Poxvirus: La característica distintiva del ADN del poxvirus es que las dos hebras de ADN se conectan. Los intermediarios replicativos, que se encuentran en el citoplasma, son concatémeros, que contienen genomas que están conectados cabeza con cabeza o cola con cola.
- Hepadnavirus: El virus de la hepatitis B utiliza la transcripción inversa para replicarse. El genoma está compuesto por un ADN circular parcialmente bicatenario, con una cadena negativa completa y una positiva incompleta. Al ingresar a la célula, la hebra positiva se completa y se transcribe. Luego, las transcripciones de ARN se transcriben inversamente en ADN mediante una enzima viral, en una serie de pasos, siguiendo de cerca el patrón de los retrovirus que incluye un salto de la nueva línea positiva a una línea repetida directamente (DR) en una dirección a otra.
Virus ARN
La replicación del genoma viral de ARN está determinada por la falta de unidades de traducción en el mensajero, que es una característica de todas las células mensajeras en animales. Para superar este problema, se han ideado tres estrategias principales.
- Este ARNm del virus funciona directamente como un mensajero que se traduce de forma monocistrónica seguido de escisión en varias proteínas.
- El ADN del virión se puede transformar para producir una variedad de ARNm monocistrónicos monocistrónicos al iniciar la transcripción en varios lugares.
- El genoma es una variedad de diferentes fragmentos de ARN que se transcriben a ARNm monocistrónico.
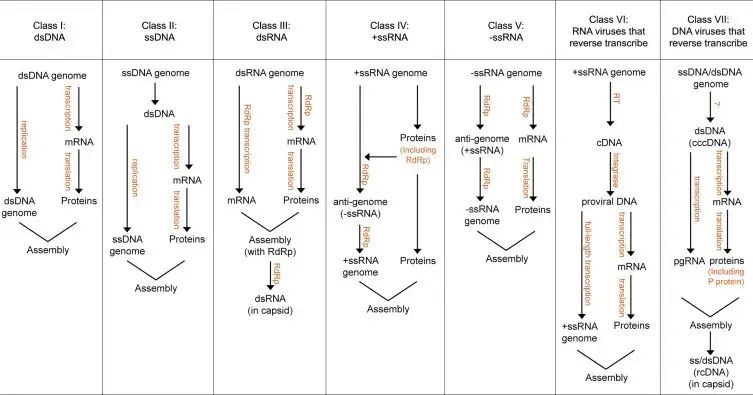
Los virus de ARN se pueden clasificar en siete clases según el ARN del virus y su relación con su mensajero.
1. Clase I (por ejemplo, picornavirus, flavivirus.)
Un genoma con polaridad +ve, actúa como mensajero, indicando información sobre la síntesis de proteínas estructurales y no estructurales.
Esta molécula de ARN también puede desencadenar la replicación que requiere primero la expresión de proteínas. Este formato proporciona poco control sobre la replicación, por ejemplo, el poliovirus no puede controlar de forma independiente el número de proteínas estructurales producidas.
2. Clase II (por ejemplo, coronavirus, togavirus.)
Muchos virus de ARN de cadena + contienen ARN subgenómico dentro del ciclo. Esto les da cierto grado de control. El ARNm subgenómico no se puede detectar a partir de la ARN polimerasa. Podría utilizarse con el único propósito de producir proteínas estructurales. Otro método para superar el problema es crear el conjunto de anidamiento de ARN. El conjunto anidado de ARN es la forma más eficaz de control. Pueden decidir la parte de su genoma que expresan.
Los togavirus son responsables de la traducción, es el caso de que el ARN de todo el genoma de 49S primero se transforma en poliproteínas que se transforman en proteínas no estructurales. Un ARNm subgenómico que se transcribe a partir del ARN de longitud completa se transforma en una poliproteína más pequeña que se transforma en proteínas estructurales del virus. Los coronavirus producen el conjunto anidado de ARNm que se generan siguiendo el mismo proceso, es decir, la transcripción -ve generada inicialmente a partir del genoma. Esta transcripción luego se transforma en dimensiones de ARNm monocistrónico monocistrónico monocistrónico. Cada uno comienza con la misma secuencia líder de 5 pies de largo que está conectada a las transcripciones cerca del comienzo de los diferentes genes y continúa hasta el extremo 3' del genoma. Estos ARNm no se pueden producir empalmando el transcrito de tamaño genómico ya que el virus puede reproducirse en células enucleadas.
3. Clase III (p. ej., paramixovirus, rabdovirus).
Es de una polaridad de -ve con respecto a su Mensajero. Una transcriptasa de ARN dependiente de ARN de virión transcribe inicialmente los genomas en señales monocistrónicas que parten de un promotor. La transcriptasa cesa y se reanuda en cada unión entre los diferentes genes.
4. Clase IV (p. ej., ortomixovirus, la mayoría de los bunyavirus).
El genoma -ve se puede encontrar en varios segmentos distintos y no superpuestos de ssRNA. La transcriptasa de virión crea un mensajero para cada pieza. Los orthomixovirus son los más comunes. Los segmentos genómicos tienen un gen, pero dos fragmentos tienen dos genes que se superponen y se expresan a través de un mensajero completo, mientras que el otro se expresa a través de un mensajero más pequeño derivado del primero a través del proceso de empalme. El proceso de replicación de los ortomixovirus es distinto de otros virus de ARN, ya que ocurre en el núcleo nuclear. La función nuclear que necesita para funcionar es a través de las tapas 5' de los mensajeros celulares que se pueden encontrar en "pellizcos" después de la escisión endonucleótica de los mensajeros de los huéspedes. Este 5′-cap se utiliza como base en la creación de mensajeros virales.
5. Clase V (por ejemplo, arenavirus, phlebovirus.)
Los arenavirus poseen un genoma ambisense. Esto significa que la mitad del genoma es de polaridad -ve y se traduce en un mensaje por la transcriptasa del virión. Sin embargo, la otra mitad que contiene +ve, se transcribe dos veces. Primero, se crea una transcripción completa del genoma y luego el ARNm se transcribe en la transcripción. Esta estrategia es evidente en la porción (pequeña) (pequeña) de los genomas de los flebovirus. Los genomas ambisense no son comunes para los virus de ARN, pero no para los virus de dsDNA.
6. Clase VI (por ejemplo, reovirus. Reovirus.)
Tienen segmentos distintos y no superpuestos de DsRNA. Cada segmento se transcribe en un ARNm que es independiente a través de la transcriptasa del virión. La mayoría de los mensajeros son monocistrónicos. Pero uno es bicistrónico y expresa otra proteína, que comienza en un AUG interno, en un marco de lectura completamente diferente. Cada segmento de ADN de reovirus se replica por separado. La transcriptasa del virión crea una nueva cadena de ARNm. Esta cadena luego se usa como plantilla para la replicasa que creará la negativa. Las dos hebras permanecen unidas a moléculas que contienen dsRNA que terminan en el virión. El proceso de replicación es asimétrico y conservador debido a que (1) la cadena -ve del virión sirve como servidor de ARN y se usa como plantilla inicial, y (2) el ARN original no se incorpora a la progenie.
7. Clase VII (p. ej., retrovirus).
Los retrovirus se diferencian de otros retrovirus en que sus genomas se han transcrito en ADN, no en ARN. Contienen dos ssRNA idénticos con polaridad +ve. Hay una cola poli A en el extremo 3′ y una tapa en el extremo 5′. Cada uno se transcribe en ADN usando transcriptasa inversa y luego se integra en el ADN de la célula, formando un provirus. La transcripción del provirus por la transcriptasa celular da como resultado la formación de moléculas virales que terminan en forma de viriones.
Debido a que los virus de ARN que pertenecen a las clases III a VII requieren transcriptasa de virión para sintetizar un mensajero, sus ARN virales purificados no son similares a los virus. Solo los de clase I, II y III están infectados. Con los virus de ARN no hay distinción entre mensajeros tardíos y tempranos.
Replicación de virus de ARN monocatenario (clases I a V)
En todos los casos, la replicación es el proceso de creación de la hebra molde que es similar a la viral de la misma longitud y que sirve como base para la progenie de hebras virales. Estos pasos son ejecutados por una variedad de enzimas, tanto de naturaleza celular como viral, junto con las nucleocápsidas que infectan a los viriones.
En muchos casos, la replicación y la transcripción pueden interferir entre sí: cuando los virus de cadena son cinco, la transcripción y la plantilla están formadas por cadenas virales. Con virus (+ve), la hebra de virus podría usarse como mediador o como plantilla de replicación. Inicialmente en el virus no hay interferencia porque la función de mensajero es suministrar las proteínas requeridas para replicarse.
En el futuro, el suministro de proteínas regulará la velocidad a la que se produce la replicación. por ejemplo, en el caso del virus de la polio, la replicación comienza cuando las proteínas pVg se unen covalentemente al extremo 5' del ARN, presumiblemente creando un complejo de replicación de ARN. Los mensajeros y su progenie frecuentemente diferían en sus características estructurales. por ejemplo, los mensajeros del virus de la gripe poseen secuencias líder con caperuza, que se derivan de mensajeros celulares.
Además, son deficientes en 17-22 nucleótidos en el extremo 3'. Además, la replicación requiere la síntesis continua de proteínas para producir las proteínas necesarias, mientras que la transcripción no.
Para la replicación del ARN, la hebra de plantilla recién creada puede permanecer unida a la hebra viral sobre la base de la cual se construye y forma una hebra de doble hebra que se extiende a lo largo del genoma viral, también conocido por el término replicativo (RF) . La síntesis de nuevas hebras se logra mediante una síntesis asimétrica que es conservadora, como los adenovirus.
Un RF que está asociado con una hebra viral de la infancia también se conoce como RI (intermedio replicativo). Las moléculas de RF son bastante abundantes en la replicación, ya que después de completarse una hebra completamente nueva, se cree que la replicasa permanece conectada durante un período de tiempo con la plantilla antes de reanudar el proceso de síntesis. Los RF se acumulan en el curso de la replicación cuando no hay más RI por formar.
Excepto los ortomixovirus. Los ARN del virus se reproducen en el citoplasma de las células. Las replicasas que están presentes en las células infectadas con el virus crean nuevas hebras de ARN viral de ambas polaridades opuestas. La transcripción ocurre en el mismo lugar donde ocurre la replicación. No está claro si la transcripción y la replicación se realizan mediante enzimas distintas oa través de una enzima.
5. Montaje
- Los virus se originan mediante la síntesis de nuevos componentes y, para que puedan escapar de las células, estos componentes deben reunirse en un lugar específico dentro de la célula y luego ensamblarse en un virus embrionario.
- De manera similar a la forma en que la penetración y la eliminación son difíciles de separar dentro del ciclo de ciertos virus, el ensamblaje a menudo puede ocurrir durante el proceso de maduración y liberación.
- El lugar de ensamblaje del virión estará determinado por el virus específico. Puede ocurrir en el núcleo de la célula o en la membrana plasmática o en varias membranas intracelulares como la del complejo de Golgi.
- La mayoría de los virus ADN que no tienen envoltura ensamblan sus nucleocápsidas en el núcleo ya que es el lugar donde se replica el genoma. Las proteínas virales se importan a través de los poros nucleares antes de llegar al sitio de ensamblaje. Después del ensamblaje, muchos virus de ADN son masivos para pasar a través de los poros nucleares, pero. A partir de ahora, ciertos virus pueden atravesar la membrana nuclear de doble membrana y algunos desencadenan la muerte celular o la apoptosis, lo que les permite salir del núcleo.
- En cambio, allí suelen agruparse los virus con envoltura que se originan en las membranas plasmáticas.
- Ácidos nucleicos en los genomas de virus como un virus helicoidal protegido por proteínas de la cápside repetidas. Esta es la razón por la que las proteínas de la cápside pueden envolver el genoma una vez que se copia el genoma (o al revés, dependiendo de cómo se conciba el virus: el genoma podría envolver las proteínas de la cápside).
- Al contrario de esto, ciertos virus icosaédricos están cerca de completar el ensamblaje de sus cápsides antes de que se agregue el genoma de ácidos nucleicos.
- El ensamblaje espontáneo de la cápside llamado "autoensamblaje" ocurre con las proteínas de la cápside de especies de virus icosaédricos simples, como picornavirus o parvovirus.
- El ensamblaje de un virus que tiene una arquitectura más compleja está controlado por una variedad de proteínas chaperonas para virus llamadas proteínas de andamiaje.
- Los adenovirus y los herpesvirus son dos ejemplos de virus icosaédricos grandes que se ensamblan con ayuda de proteínas de andamiaje.
6. Maduración
- Una vez que el genoma del ácido nucleico, así como otras proteínas, están contenidos dentro de la cápside, se construye a partir de una o más proteínas virales de traducción. Los próximos pasos de la replicación tienen lugar la maduración y liberación.
- En este punto, el virión se desarrolla como célula. Si la célula se abriera después de este tiempo, el virión no estaría en condiciones de comenzar a infectar nuevas células.
- La maduración se refiere específicamente a los cambios finales en un virión inmaduro, que dan como resultado la formación de una partícula viral infecciosa. Los cambios estructurales de la cápside típicamente están involucrados y están mediados por enzimas del huésped o enzimas codificadas por virus.
Mecanismo de maduración
Influenza Virus
Un gran ejemplo es que uno de ellos es la proteína H de la influenza. Está involucrado en la unión con los ácidos siálicos de las células, como se mencionó anteriormente, y la proteína tiene la capacidad de unirse al ácido siálico después de haber sido glicosilada (a través de modificaciones postraduccionales). La proteína debe descomponerse en dos partes que son HA1 y la otra para que se vuelva infecciosa. Esto se debe a que, aunque la porción HA1 puede unirse a los receptores de la superficie de las células, la otra parte es la que une la envoltura del virus a la membrana de la célula endosómica para liberar virus en el citoplasma de las células. La escisión de la proteína HA en HA1 y HA2 se produce a través de proteasas celulares (enzimas que escinden proteínas).
VIH
Contrariamente a esto, la partícula central del VIH está formada por las proteínas codificadas por los genes gag. El gen se transforma en una poliproteína que se descompone a través de la proteasa del VIH para crear las proteínas de la matriz, la cápside y la nucleocápside que componen el virión. En este escenario, la maduración tiene lugar cuando el virión se elimina de la superficie celular y es necesaria para la formación de un virión infeccioso. En el Capítulo 8, “Vacunas, antivirales y los usos beneficiosos de los virus”, muchos medicamentos contra el VIH funcionan evitando las acciones de la proteasa del VIH, lo que evita la escisión de la proteína y la subsiguiente creación del virión infeccioso.
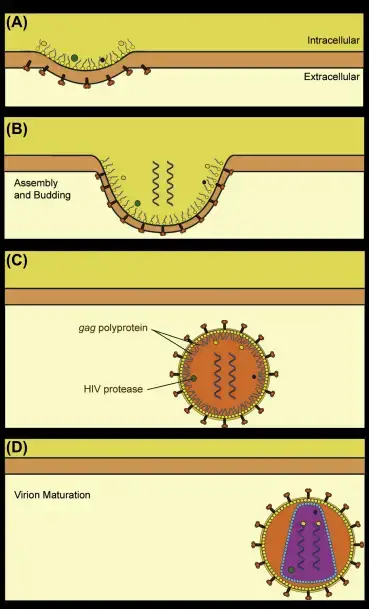
El ensamblaje, maduración y liberación de HIV Violons se produce como resultado de los siguientes pasos:
- (A) Las proteínas del VIH se congregan en las membranas plasmáticas de las células y hacen que se desarrolle un brote en la membrana.
- El ADN del genoma diploide se empaqueta dentro de la cápside que se ensambla (B).
- El virus se ha dispersado por la membrana de las células, sin embargo, la poliproteína Gag no se ha descompuesto para separar las proteínas de la matriz y la cápside del virión (C).
- La proteasa del VIH corta la poliproteína y permite que las proteínas construyan la estructura del virión (D).
7. Lanzamiento
- La etapa final en el ciclo de replicación es la liberación del virión al entorno circundante para continuar infectando liberando nuevas células.
Mecanismos involucrados en la liberación
La liberación puede ocurrir de muchas maneras diferentes, según el virus.
En ciernes
- Los virus que obtienen sus envolturas de la membrana plasmática generalmente pueden formar una envoltura en la capa interna de la membrana plasmática, encapsulando sus proteínas de envoltura en la membrana plasmática.
- Cuando las proteínas de la cápside del virus están en contacto con las proteínas virales asociadas a la membrana, crean una membrana plasmática que comienza a curvarse sobre la cápside.
- El proceso continúa hasta que el plasma se ha encerrado completamente alrededor del virus y es cuando la célula deja de existir. Esto se llama brotar.
- El virus puede desarrollarse a partir de cualquier membrana de la célula, como el sistema rER, el complejo de Golgi e incluso la envoltura nuclear.
Exocitosis
- En este caso, el virión que ya está envuelto no requiere moverse a través de la membrana plasmática. Por lo general, se exocitosis para soltar la célula.
- El virus sin envoltura también puede salir de la célula por exocitosis.
Lisis
- Los virus líticos provocan la alteración de la membrana plasmática y desencadenan la lisis o la ruptura de las células. Luego, el virus libera su forma naciente para infectar las nuevas células.
- Los virus humanos que no están envueltos se liberan a través de la lisis celular.
Referencias
- Louten J. Virus Replication. Essential Human Virology. 2016:49–70. doi: 10.1016/B978-0-12-800947-5.00004-1. Epub 2016 May 6. PMCID: PMC7149683.
- https://virology-online.com/general/Replication.htm
- https://en.wikipedia.org/wiki/Viral_replication
- https://www.immunology.org/public-information/bitesized-immunology/pathogens-and-disease/virus-replication
- https://courses.lumenlearning.com/boundless-microbiology/chapter/viral-replication/
- https://www.news-medical.net/health/How-does-Viral-Replication-Work.aspx
- https://www.sciencedirect.com/topics/medicine-and-dentistry/virus-replication
- https://www.frontiersin.org/articles/10.3389/fmicb.2018.01546/full