
Abstract
Background and aims
Naturalized alien and native plants can impact each other directly when they grow next to each other, but also indirectly through their soil legacies. These alien-native interactions can also be modified by the presence of a third alien or native species. However, it is unknown how the performance of co-growing alien and native species is affected by their soil legacies and by the presence of an additional species.
Methods
In our two-phase plant-soil-feedback experiment, soils were first conditioned by eight herbaceous species, four of which are naturalized and four of which are native to Germany. We then grew all 16 pairwise alien-native species combinations on soil conditioned by the respective alien or native species, on a mixture of soils conditioned by both species or on control soil. Each pair of test plants was grown on these soils without or with an additional alien or native species.
Results
Soil conditioning, and particularly conspecific soil conditioning, reduced growth of the alien and native test plants. The addition of another species also reduced growth of the test plants. However, the negative conspecific soil-legacy effect on alien test plants was reduced when the additional species was also alien.
Conclusion
The negative conspecific plant-soil feedback for alien and native plants in our study could promote their coexistence. However, as partial alleviation of negative conspecific effects on alien plants occurred with an additional alien species, the chances of coexistence of alien and native species might decrease when there are multiple alien species present.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Humans have accidentally or intentionally transported and introduced thousands of species from their native regions into regions novel to those alien species (Hulme et al. 2008). The number of alien species that have now established self-sustaining populations in the wild (i.e. have become naturalized) has dramatically increased over the past centuries (Dawson et al. 2017; Seebens et al. 2017). For plants, already more than 13,000 species have become naturalized (van Kleunen et al. 2015), and this number is likely to continue to increase (Seebens et al. 2021). Plant invasions result in a global floristic homogenization (Yang et al. 2021), and may threaten native species (Pouteau et al. 2023; Vilà et al. 2011). Therefore, many studies have focused on how alien and native species interact and whether they can coexist. Moreover, as more and more alien species naturalize and increase in dominance, alien species are increasingly likely to not only interact with native species but also with other naturalized alien species. Therefore, an important question is how alien and native co-growing plant species and their interactions are affected by the presence of other alien plant species.
Plant-soil interactions play a major role in determining plant growth and species coexistence (Callaway et al. 2004; Fahey et al. 2020; Wolfe and Klironomos 2005). Plants alter the biotic and abiotic properties of the soil, and thereby they leave a soil legacy that can affect the performance of subsequent plants, a phenomenon frequently referred to as plant-soil feedback (Bever et al. 1997; van der Putten et al. 2013). Soil-legacy effects are frequently negative, and those of conspecifics are usually more negative than those of heterospecifics, most likely due to the accumulation of species-specific enemies. When conspecific plant-soil feedback is more negative than heterospecific plant-soil feedback, this should result in negative frequency-dependent population dynamics that favour species coexistence (Mangan et al. 2010). However, because alien species are likely to be released from some of their specialist root herbivores and pathogens (Blumenthal et al. 2009; Keane and Crawley 2002; Mitchell and Power 2003), they may be less negatively affected by soil legacies than native species. Furthermore, secondary metabolites, such as those in root exudates, of species can also shape their soil legacies, and it is thought that the novelty of these chemicals produced by alien species can allow them to become dominant over native plants (Callaway and Ridenour 2004). While multiple studies have found evidence that conspecific plant-soil feedback effects are less negative for alien than for native species (Dostálek et al. 2016; Van Grunsven et al. 2007), this is not always the case (Chen et al. 2024; Meisner et al. 2014), and most of these studies tested the effects on single plants and not on co-growing alien and native species.
Through their soil legacies, alien plant species can benefit the growth of other alien plant species that arrive later (Chen and van Kleunen 2022). Moreover, differences in soil-legacy effects of alien species on alien and native species that subsequently grow on the soil could drive the outcome of competition (Zhang et al. 2020b). So, when an additional alien plant species enters a native community that includes already another alien, its growth might indirectly be promoted, which would be in line with the predictions of the invasional meltdown hypothesis (Simberloff and Von Holle 1999). However, as many naturalized alien plants, at least the invasive ones, usually have a high competitive ability, the addition of another alien species could also result in a reduced growth of the alien species that was already present due to more intense competition (Haeuser et al. 2019). Therefore, how a third species generally influences soil-legacy and competitive effects remains unknown.
Here, we conducted a two-phase plant-soil-feedback experiment to test whether and how soil legacies of alien and native plants, and the presence of an additional species affect the growth of co-growing alien and native species. We first grew eight herbaceous species, four of which are naturalized and four of which are native to Germany, separately in pots to condition the soil. In the second phase, we then grew all 16 pairwise alien-native species combinations on unconditioned control soil or soils conditioned by the respective alien species, the respective native species or a mixture of both, and with or without an additional alien or native species. We tested whether the growth of the alien and native test species was affected by (1) the different soil legacies, (2) the presence of an additional alien or native species, and (3) the interaction between soil legacies and the presence of an additional species.
Materials and methods
Study species and seed material
As study species (Table S1), we selected four herbaceous plant species that are widely naturalized aliens in Germany (Bidens frondosa L., Lepidium virginicum L., Lolium multiflorum Lam. and Solidago canadensis L.) and four that are widely distributed natives (Bromus sterilis L., Centaurea jacea L., Daucus carota L. and Plantago lanceolata L.). The eight species are typically found in ruderal or grassland habitats (FloraWeb database; www.floraweb.de, accessed March 2021), and are from five families (Table S1). Seeds of three of the eight species were ordered from Rieger-Hofmann GmbH, and seeds of the other five species came from the seed collection of the Botanical Garden of the University of Konstanz (Table S1).
Experimental set up
Soil-conditioning phase
From 12 to 21 April 2021, we sowed each of the eight species for the soil-conditioning phase separately into trays (18 cm × 14 cm × 5 cm) filled with potting soil (Topferde; Einheitserde Co., Sinntal-Altengronau, Germany). To make sure that all seedlings would be in a similar developmental stage at transplanting, seeds were sown on different dates (Table S1) based on prior knowledge about the time required for germination. The pre-cultivation was done in a greenhouse of the Botanical Garden of the University of Konstanz, Germany (47°41′32″N, 9°10′41″E), and the temperature was maintained between 18 and 25 °C.
To make sure that the substrate used in the soil-conditioning phase contained live soil organisms, on 26 April 2021, we collected field soil from a native grassland site near the greenhouse where at least three of the four native study species but none of the alien study species occurred (Table S7). To remove large pebbles and plant fragments, we sieved the field soil using a metal grid with a mesh width of 1 cm. We then filled 2-L pots (14 cm × 14 cm × 14.5 cm) with a substrate consisting of a mixture of 25% field soil, 37.5% sand and 37.5% vermiculite (by volume). We used sand and vermiculite to facilitate removal of the roots at the end of the conditioning phase. On 3 May 2021, we transplanted one seedling into the center of each pot. To obtain sufficient amounts of conditioned soil, we had 96 replicate pots for each of the eight species (i.e. 768 pots). In addition, we had 256 pots of substrate without any plant as a control treatment, resulting in a total of 1024 pots in the soil-conditioning phase. Seedlings that died within two weeks after transplanting were replaced. All pots were individually placed on plastic saucers (Ø = 17 cm) and randomly allocated to positions in three greenhouse compartments (24 °C/18°C day/night temperature, 16 h/8 h day/night). We watered all pots every 2–3 days, and fertilized them four times (13 May, 30 May, 18 June and 7 July 2021) with a water-soluble fertilizer (1‰ m/v, Universol Blue with an NPK ratio of 3:2:3) to reduce negative effects due to nutrient depletion in the next phase (Brinkman et al. 2010). To reduce potential effects of environmental heterogeneity within and among the greenhouse compartments, we re-randomized the positions of the pots five weeks after the start of the soil-conditioning phase.
As it is generally thought that the soil-conditioning phase should last at least two months (van der Heijden 2004), we harvested the plants and soils on 19 July 2021 (11 weeks after the start of the soil-conditioning phase). The soil of each pot was sieved through a 5-mm mesh to remove the roots. After each pot, the mesh was sterilized with 70% ethanol. The sieved soil was stored at 4 °C until use in the test phase.
Test phase
From 12 to 19 July 2021, we sowed the eight species to produce seedlings for the test phase (Table S1). The pre-cultivation conditions were the same as for the soil-conditioning phase. We filled 0.5-L pots (9 cm × 9 cm × 8 cm) with soil from the conditioning phase. To test the effects of the variously conditioned soils on each of the 16 alien-native species pairs, we used four soil-conditioning treatments in the test phase (Fig. 1): (1) soil conditioned without a plant (Control), (2) soil conditioned by the alien species of the respective species pair (Alien), (3) soil conditioned by the native species of the respective species pair (Native), and (4) a 1:1 mixture of soils conditioned by the alien and native species of the respective pair (Mixed). So, in the Mixed treatment, we mixed soil from one pot conditioned by the alien species and from one pot conditioned by the native species. Alternatively, we could have included pots with both the alien and native species in the conditioning phase, but then each of those pots would have had one more individual than the single-species pots. As mixing soils from different pots could increase the diversity of soil biota (Reinhart and Rinella 2016; Rinella and Reinhart 2018), we also mixed soils from two soil-conditioning-phase pots for the Control, Alien and Native treatments. For example, for a test-phase pot in the Alien treatment of the species pair Solidago canadensis - Bromus sterilis, we mixed soils conditioned by S. canadensis from two pots. Zhang et al. (2020a) found that such soil mixing did not affect performance of the test-phase plants. Each of the soil-conditioning-phase pots, including the controls, was used only once, so that all the pots in the test phase were independent from one another.

Overview of the experimental design. In the soil-conditioning phase (from 3 May to 19 July 2021), the soil was conditioned separately by each of the four alien and four native species, or without any plants. In the test phase (from 2 August to 4 October 2021), pots were created for each of four soil-legacy treatments by filling them with soil without plant conditioning (Control), soil conditioned by an alien species (Alien), soil conditioned by a native species (Native), or a mixture of soil conditioned by an alien species and soil conditioned by a native species (Mixed). The alien and native test plants that were planted corresponded to the species that had been used to condition the soil. For pots in the additional species treatment, a plant of one of the other six alien or native species was also planted
On 2–3 August 2021, we transplanted seedlings into the test-phase pots. For each of the 16 alien-native species pairs, we had 32 pots; eight for each of the four soil-conditioning treatments (Control, Alien, Native, Mixed). In each of these pots, we planted one seedling of each of the two species. Six of the eight pots per soil-conditioning treatment of a species pair received one additional seedling (i.e. a third plant) of one of the other six species (3 native and 3 alien species). Each of those other species was used only once per soil-conditioning treatment of a species pair, so that we effectively had three replicates (n = 3) with an additional alien species and three replicates with an additional native species. The two remaining pots per soil-conditioning treatment of a species pair served as controls for the effects of the presence of an additional species (n = 2). In these pots without additional species, the two seedlings were planted 6 cm apart: 3 cm to the left and 3 cm to the right of the center of the pot. In the pots with an additional species, the three seedlings were also planted 6 cm apart but in a centrosymmetric equilateral triangle. In total, we had 512 pots in the test phase.
All pots were individually placed on plastic saucers (Ø = 15 cm) and randomly allocated to positions in a greenhouse compartment (24 °C/18°C day/night temperature, 16 h/8 h day/night). We watered the pots every 2–3 days and fertilized them two times (16 August and 13 September 2021) with 80 mL of a water-soluble fertilizer (1‰ m/v, Universol Blue). Positions of all pots were re-randomized four weeks after the start of the test phase.
Measurements
On 4–5 August 2021, at the start of the test phase, we measured the length and width of the largest leaf on each seedling, and counted the number of leaves. The initial size of each seedling was then estimated as length of the largest leaf × width of largest leaf × number of leaves. On 4 October 2021, nine weeks after the start of the test phase, we harvested the aboveground biomass of each plant separately. Because it was impossible to separate the roots of the different species in each pot, the belowground biomass was not harvested. The plant materials from both the soil-conditioning and test phases were dried at 70 °C to constant weight, and were then weighed.
Statistical analysis
To test whether the soil-legacy and the additional-species treatments affected the performance of the alien and native species in the test phase, we fitted a linear mixed effect model with the lme function in the R package ‘nlme’ (Pinheiro et al. 2019). Biomass of each individual test plant (i.e. one native and one alien plant per pot) was included as the response variable. The origin of each individual test plant (alien, native), soil-legacy treatment (Control, Alien, Native, Mixed), the additional-plant treatment (without, with alien, with native) and their interactions were included as fixed effects. We additionally ran three orthogonal hierarchical contrasts (Casella 2008) for the soil-legacy treatments (SL). First, to test the average effect of soil conditioning by a plant, we compared Control vs. the average of Alien, Native and Mixed (SLControl/Conditioned). Second, to test whether the effects of soils conditioned by both species of a species pair differ from those conditioned by only one of the two species, we compared Mixed vs. the average of Alien and Native (SLMixed/Single). Third, to test whether the effects of soils conditioned by the alien species of a species pair differed from those conditioned by the native species, we compared Alien vs. Native (SLAlien/Native). We also ran two orthogonal hierarchical contrasts for the additional-plant treatment (AP). First, to test the average effect of the presence of an additional species, we compared the treatment without an additional species vs the average of the treatments with an additional alien or native species (APWithout/With). Second, to test the effect of the status of the additional species, we compared the treatment with an additional alien species vs. the treatment with an additional native species (APAlien/Native). To account for nonindependence of test species belonging to the same family and of plants belonging to the same test species, family and species identity of the test plant were used as random effects. To account for the identity of the additional species, it was also included as a random effect. Furthermore, to account for nonindependence of the two test plants (one alien and one native) that were in the same pot, identity of the test-phase pot was also included as a random effect. Finally, to account for differences in initial sizes of the test plants, we included our estimates of initial size as a fixed covariate.
In addition to analysing the individual biomass of each alien and native test plant, we also tested whether the soil-legacy and additional-species treatments affected the biomass proportion of the alien test plants in the pots. Therefore, we fitted a model including the proportional biomass of the alien test plant in each pot, calculated as biomassalien/(biomassalien + biomassnative), as a response variable. We included the soil-legacy and additional-species treatments and their interaction as fixed effects. Species identities of the alien, native and additional plants were included as random effects, and proportional initial size of the alien test plant was included as a covariate. In addition, to test whether the total aboveground biomass (also including the third plant, if present) per pot was affected by the treatments, we ran a similar model for total biomass per pot. Here, we used the combined initial size estimates of all plants in a pot as a covariate. Furthermore, because the soil-condition effect on plants in the test phase might depend on the biomass produced by the plants that conditioned the soil (as e.g. larger plants might have depleted more nutrients), we also ran a model for total aboveground biomass per pot after excluding the Control treatment. In this model, the average aboveground biomass of the two conditioning plants used to produce the soil in the respective test-phase pot (for soil mixing) was added as a covariate.
To additionally test whether the soil-legacy treatment affected the additional plant, we also fitted a model using the subset of pots with additional plants. In this model, we included proportional biomass of the additional plant, calculated as biomassadditional/(biomassadditional + biomassalien + biomassnative), as the response variable. We included the origin of the additional species (alien or native), soil-legacy treatment (included as three orthogonal hierarchical contrasts: SLControl/Conditioned, SLMixed/Single and SLAlien/Native) and their interactions as fixed effects. Species identities of the alien, native and additional plants were included as random effects, and proportional initial size of the additional species was included as a covariate.
In all models, to improve homoscedasticity of residuals, we allowed the variance to vary among the test species and among the additional species by using the varIdent and varComb functions (Table S6). Furthermore, to meet the assumption of normality of the residuals, biomass of the test plant, biomass proportion of alien test plant and biomass proportion of the additional plant were square-root transformed. We used log-likelihood ratio tests to assess significance of the fixed effects by comparing models with and without the effect of interest (Zuur et al. 2009). All analyses were conducted in R 4.2.2 (R Core Team 2019).
Results
Effects of soil legacies and additional plants on test plants
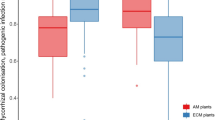
On average, the aboveground biomass of alien and native test species, as well as the total biomass per pot, were reduced when plants were grown on soil conditioned by plants instead of on control soil not conditioned by any plant (Tables 1 and S2, Figs. 2 and S1). The individual test plants achieved more biomass in the Mixed soil treatment compared to the average of the Alien and Native soil treatments when there was no additional competitor present (+ 14.1%), whereas the reverse was true in the presence of an additional competitor (-7.3%; significant APWithout/With × SLMixed/Single in Table 1; Figs. 2 and S4). Furthermore, conspecific plant-soil feedback was more negative than heterospecific plant-soil feedback, as the alien test plants produced the least biomass on soil conditioned by the alien species, and the native test plants produced the least biomass on soil conditioned by the native species (significant O × SLAlien/Native interaction in Table 1; Fig. 2). As a consequence, although the alien test plants produced more biomass than the native plants in most soil-conditioning treatments, this difference almost disappeared when the plants were grown on soils conditioned by the alien species. The presence of an additional plant reduced the biomass of the test species. However, the magnitude of this reduction depended on the alien/native origins of the additional plant, the test plant and the soil-conditioning species (significant O × APAlien/Native × SLAlien/Native interaction in Table 1; Figs. 2 and S5). More specifically, the negative conspecific soil-legacy effect on alien test plants was partly alleviated when an additional alien species instead of an additional native species was present (Figs. 2 and S5). Consequently, the difference in biomass between alien and native test plants on soil conditioned by the alien species was smaller when the additional plant was native instead of alien.

Effects of the presence of an additional species on the aboveground biomass of alien and native test plants in the four soil-legacy treatments. Shown are modelled means (± SEs). Density ridgelines indicate the distribution of modelled values. The underlying statistical model is shown in Table 1, and significant contrasts are visualized in Fig. S4
Effects of soil legacies and additional species on proportional biomass of alien plants
The proportional biomass of the alien test plant per pot (biomassalien/[biomassalien + biomassnative]) was lowest when grown on soil conditioned by the alien species and highest on soil conditioned by the native species (Table 2; Fig. 3). However, this difference in proportional biomass of the alien test plant between the alien and native soil-legacy treatments tended to be reduced by the presence of an additional alien (marginally significant APAlien/Native × SLAlien/Native interaction in Table 2; Fig. 3). In other words, the presence of an additional alien instead of an additional native species improved the biomass proportion of the alien test plant when grown on soil conditioned by the alien species (+ 7.5%) and decreased it when grown on soil conditioned by the native species (-8.2%). As a consequence, alien test plants experienced less negative conspecific feedback effects when grown with an additional alien species.

Effects of the presence of an additional species on the biomass proportion of alien test plants in four soil-legacy treatments. Proportional biomass of alien test plants was calculated as biomassalien/(biomassalien + biomassnative). Shown are modelled means (± SEs). Density ridgelines indicate the distribution of modelled values. Colors indicate the treatment with the absence (grey) and the presence of alien (orange) or native (purple) additional species. The underlying statistical model is shown in Table 2
Discussion
We tested whether and how the performance of co-growing alien and native plants was affected by soil legacies of one or both of the two species and by the presence of an additional (i.e. a third) alien or native species. As expected, we found that plants produced less biomass when the soil had been conditioned by a plant instead of without one, indicating negative plant-soil feedback. Moreover, plants experienced more negative plant-soil feedback when the soil had been conditioned by a conspecific instead of a heterospecific plant. Consequently, although alien plants in our study produced overall more biomass than the co-growing native ones, this difference was reduced when the soil had been conditioned by the alien instead of the native species. This indicates that alien plants, just like the native ones, can also suffer negative conspecific feedback effects. Interestingly, although the presence of an additional species resulted in a reduction of the biomass of the two test plants, indicating competition, proportional biomass of the alien test plants, relative to the joint biomass of the two test plants, on soil conditioned by a conspecific was slightly increased when there was another alien species present. This suggests that the presence of another alien species partly alleviated the negative conspecific plant-soil-feedback effects on alien plants.
In line with the results of many previous studies (Crawford et al. 2019; Lekberg et al. 2018), we found that the test plants experienced negative plant-soil feedback effects on soils that had been conditioned by a previous plant (Fig. 2). This could reflect that the conditioned soils contained less nutrients than the control soils. However, although the biomass produced on the soil during the conditioning phase had a negative effect on biomass production per pot in the test phase, this effect was relatively weak (Table S3, Fig. S3), most likely because we had fertilized the pots during the conditioning and test phases. Furthermore, if solely nutrient depletion would explain the effect of soil conditioning, then the test plants and the additional plant in a pot should be similarly affected. However, the additional plants had increased their proportional biomass on the conditioned soils (Table S5, Fig. S2b). Therefore, the negative plant-soil feedback is most likely predominantly caused by the accumulation of herbivorous and pathogenic soil organisms.
Most previous studies found that biomass production is lower on soils conditioned by conspecifics than on soil conditioned by heterospecifics (e.g. Lozano et al. 2022; MacDougall et al. 2011; Rutten et al. 2016). This was also the case in our study, as both aliens and natives had the lowest biomass on soils conditioned by conspecifics. However, the conspecific plant-soil feedback was less negative for the alien plants (-39.5%) than for the native plants (-48.5%; Fig. 2). This could reflect that the soil inoculum used in the conditioning phase came from a site in which the majority of our native study species occurred naturally (Table S7). This field soil was used to increase the chance that specialist herbivores and pathogens of the native test species would be present, and to avoid that the results would be dominated by generalists that would attack both the native and alien species equally. Our results are thus in line with the predictions of the enemy-release hypothesis, i.e. that alien species have been released from some of their species-specific natural enemies (Keane and Crawley 2002; Klironomos 2002; MacDougall et al. 2011; Mitchell and Power 2003).
Although averaged across the three soils conditioned by plants (i.e. Alien, Native, Mixed) the biomass of test plants was lower than when grown on the control soil, the biomass production of test plants on the soils conditioned by heterospecifics was actually not much lower than on the control soils (Fig. 2). This may suggest that there was no heterospecific plant-soil feedback. However, in our experimental setup, soil that was heterospecific for one of the two test plants in a pot was conspecific for the other one. Therefore, if one of the two test plants in a pot suffered from negative conspecific plant-soil feedback, the other test plant in the same pot most likely benefited from the reduced growth of the other test plant (Lekberg et al. 2018). This indirect effect might have compensated any direct negative heterospecific plant-soil feedback effects in our experiment.
Like in many other studies (Vilà and Weiner 2004; Wang et al. 2017), we found that the alien plants produced on average more biomass than the native plants (Fig. 2). The inherently faster growth of many naturalized alien plants may increase their competitive ability (Zhang and van Kleunen 2019), and thus may contribute to their invasion success. Here, however, we found that the negative conspecific plant-soil feedback resulted in a smaller biomass difference between the alien and native plants on the soils conditioned by the alien species (Fig. 2). Consequently, the proportional biomass of the alien plants was lowest on soils conditioned by the alien species and highest on soils conditioned by the native species (Fig. 3). This finding suggests that the advantage that the alien species may have over the native species when they start to invade a community —and when the soil is thus conditioned by natives only— might disappear over time, increasing the likelihood of coexistence.
On soils conditioned by both the alien and native species (i.e. the Mixed treatment), the proportional biomass of the alien plant was intermediate, as expected. This was the case both with and without additional species, but, without additional species, the biomass of the test plants in the mixed soils exceeded the average of the biomass on each of the separate soils (+ 14.1%; Fig. S4). This suggests that the test plants experienced relatively weak negative plant-soil feedbacks on mixed soils. It has been suggested that the mixing of soils conditioned by different species can increase the diversity of soil legacies and thus increase the probabilities that the test plant will interact with them (Thakur et al. 2021; van der Heijden et al. 2008). However, species-specific pathogens and secondary metabolites, such as allelochemicals, in the soils will also be diluted during this process, and the same applies if each species would have depleted different specific nutrients. These dilution effects should reduce the negative effects of the mixed soils on plant growth. In addition, soil physical and chemical properties, such as soil pH and electric conductivity, could also change during the formation of soil legacies. Future studies on plant-soil feedbacks should consider these soil properties.
When an increase in the number of species coincides with an increase in plant density, as was the case in our experiment, this usually results in more intense competition (Grace and Tilman 1990; Weigelt and Jolliffe 2003). Consistent with this, we found that growth of the alien and native test plants was reduced in the presence of the additional (i.e. third) species. Although the alien plants in our study were generally larger than the native ones, the effect of the additional species did not significantly depend on its origin. This suggests that in our study, the intensity of competition was more strongly affected by the density of individuals than by the origin of the additional species. However, the reduction in biomass advantage of the alien over the native test plants on soils conditioned by the alien species was smaller when the additional species was an alien instead of a native species (Figs. 3 and S5). Consequently, the proportional biomass of the alien test plant, when grown on soils conditioned by a conspecific, was highest when the additional species was also an alien. Plants usually experience stronger competition from tall than from small plants (Dostál 2011; Feng et al. 2016). Therefore, a possible explanation for our finding could be that the native test species, which were overall smaller than the alien species, suffered more from the additional alien competitor than the alien test species did. This is in line with results of a meta-analysis that invasive alien plants had stronger negative effects on natives than on aliens (Kuebbing and Nuñez 2016). So, while overall, the presence of an additional alien species had a direct negative effect on the biomass of the alien test plants, it had indirectly —through its negative effect on the native test plants— a positive effect of the alien test plants. Such a positive indirect effect is one of the possible mechanisms that may underlie invasional meltdown (Simberloff and Von Holle 1999).
Although the alien test plants, when grown on conspecific soil, benefited from the presence of another alien species, the additional alien species neither benefited nor incurred a cost from growth on soil conditioned by the alien test species (Tables S4-S5, Fig. S2). Invasional meltdown, however, does not require that the alien species are mutually beneficial to one another (Simberloff and Von Holle 1999). Indeed, unidirectional positive effects for just one of the alien species are common (e.g. Flory and Bauer 2014; Jäger et al. 2009; Relva et al. 2010). As plant-soil-feedback-mediated invasional meltdown is a temporal one-way process (i.e. from earlier to later in time), it is more likely that only the subsequent alien species benefits from the soil legacies of a previous alien species. Indeed, Chen and van Kleunen (2022) found that subsequent alien plants experienced less negative soil-legacy effects on soils conditioned by heterospecific alien plants. Furthermore, Zhang et al. (2020b) found that the soil legacies of alien plants benefited later alien plants when growing in competition with native plants. In our study, although the additional alien plants did not benefit from the soil conditioned by the other alien plant (i.e. the test alien plant), it was not at a competitive disadvantage either. As a consequence, despite the combined effects of competition and soil legacies, the subsequent alien plants, irrespective of whether they were test or additional plants, had overall an advantage over the native plants. Exploring the specific mechanisms behind this would be worthy of consideration in future research.
In conclusion, we found that soil legacies and the presence of an additional species negatively affected the biomass of co-growing native and alien plants. In the absence of an additional species, test plants grew better on soil conditioned by both test species than on soils conditioned by only the native or the alien test species. Moreover, both alien and native plants grew worse on soil conditioned by conspecifics than on soils conditioned by heterospecifics, and native plants were more negatively affected by this than alien plants. The negative conspecific plant-soil feedback of the alien plants might promote coexistence (van der Putten et al. 2013), and thereby prevent the alien plants from driving the native plants to extinction. However, as the negative conspecific plant-soil feedback experienced by alien test plants was partly alleviated by the presence of another alien species, the chances of coexistence of the alien and native species might decrease when there are multiple alien species present.
Data availability
The raw data are available on request to the corresponding author.
References
Bever JD, Westover KM, Antonovics J (1997) Incorporating the soil community into plant population dynamics: the utility of the feedback approach. J Ecol 85:561–573
Blumenthal D, Mitchell CE, Pyšek P, Jarošík V (2009) Synergy between pathogen release and resource availability in plant invasion. Proc Natl Acad Sci USA 106:7899–7904
Brinkman EP, Van der Putten WH, Bakker EJ, Verhoeven KJF (2010) Plant-soil feedback: experimental approaches, statistical analyses and ecological interpretations. J Ecol 98:1063–1073
Callaway RM, Ridenour WM (2004) Novel weapons: invasive success and the evolution of increased competitive ability. Front Ecol Environ 2:436–443
Callaway RM, Thelen GC, Rodriguez A, Holben WE (2004) Soil biota and exotic plant invasion. Nature 427:731–733
Casella G (2008) Statistical design. Springer, New York, NY
Chen D, van Kleunen M (2022) Invasional meltdown mediated by plant-soil feedbacks may depend on community diversity. New Phytol 235:1589–1598
Chen D, van Kleunen M, Wang YJ, Yu FH (2024) Invasive and native plants show different root responses to feedback-mediated soil heterogeneity. Plant Soil 494:497–508
Crawford KM, Bauer JT, Comita LS, Eppinga MB, Johnson DJ, Mangan SA, Queenborough SA, Strand AE, Suding KN, Umbanhowar J, Bever JD (2019) When and where plant-soil feedback may promote plant coexistence: a meta-analysis. Ecol Lett 22:1274–1284
Dawson W, Moser D, van Kleunen M, Kreft H, Pergl J, Pyšek P, Weigelt P, Winter M, Lenzner B, Blackburn TM, Dyer EE, Cassey P, Scrivens SL, Economo EP, Guénard B, Capinha C, Seebens H, García-Díaz P, Nentwig W, García-Berthou E, Casal C, Mandrak NE, Fuller P, Meyer C, Essl F (2017) Global hotspots and correlates of alien species richness across taxonomic groups. Nat Ecol Evol 1:0186
Dostál P (2011) Plant competitive interactions and invasiveness: searching for the effects of phylogenetic relatedness and origin on competition intensity. Am Nat 177:655–667
Dostálek T, Münzbergová Z, Kladivová A, Macel M (2016) Plant–soil feedback in native vs. invasive populations of a range expanding plant. Plant Soil 399:209–220
Fahey C, Koyama A, Antunes PM, Dunfield K, Flory SL (2020) Plant communities mediate the interactive effects of invasion and drought on soil microbial communities. ISME J 14:1396–1409
Feng Y, van Kleunen M, Huenneke L (2016) Phylogenetic and functional mechanisms of direct and indirect interactions among alien and native plants. J Ecol 104:1136–1148
Flory SL, Bauer JT (2014) Experimental evidence for indirect facilitation among invasive plants. J Ecol 102:12–18
Grace JB, Tilman D (1990) Perspectives on plant competition. Academic Press, San Diego
Haeuser E, Dawson W, van Kleunen M (2019) Introduced garden plants are strong competitors of native and alien residents under simulated climate change. J Ecol 107:1328–1342
Hulme PE, Bacher S, Kenis M, Klotz S, Kühn I, Minchin D, Nentwig W, Olenin S, Panov V, Pergl J, Pysek P, Roques A, Sol D, Solarz W, Vilá M (2008) Grasping at the routes of biological invasions:: a framework for integrating pathways into policy. J Appl Ecol 45:403–414
Jäger H, Kowarik I, Tye A (2009) Destruction without extinction: long-term impacts of an invasive tree species on Galapagos highland vegetation. J Ecol 97:1252–1263
Keane RM, Crawley MJ (2002) Exotic plant invasions and the enemy release hypothesis. Trends Ecol Evol 17:164–170
Klironomos JN (2002) Feedback with soil biota contributes to plant rarity and invasiveness in communities. Nature 417:67–70
Kuebbing SE, Nuñez MA (2016) Invasive non-native plants have a greater effect on neighbouring natives than other non-natives. Nat Plants 2:16134
Lekberg Y, Bever JD, Bunn RA, Callaway RM, Hart MM, Kivlin SN, Klironomos J, Larkin BG, Maron JL, Reinhart KO, Remke M, van der Putten WH (2018) Relative importance of competition and plant-soil feedback, their synergy, context dependency and implications for coexistence. Ecol Lett 21:1268–1281
Lozano YM, Aguilar-Trigueros CA, Ospina JM, Rillig MC (2022) Drought legacy effects on root morphological traits and plant biomass via soil biota feedback. New Phytol 236:222–234
MacDougall AS, Rillig MC, Klironomos JN (2011) Weak conspecific feedbacks and exotic dominance in a species-rich savannah. Proc Roy Soc B-Biol Sci 278:2939–2945
Mangan SA, Schnitzer SA, Herre EA, Mack KML, Valencia MC, Sanchez EI, Bever JD (2010) Negative plant-soil feedback predicts tree-species relative abundance in a tropical forest. Nature 466:752–755
Meisner A, Hol WHG, de Boer W, Krumins JA, Wardle DA, van der Putten WH (2014) Plant-soil feedbacks of exotic plant species across life forms: a meta-analysis. Biol Invasions 16:2551–2561
Mitchell CE, Power AG (2003) Release of invasive plants from fungal and viral pathogens. Nature 421:625–627
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team (2019) nlme: linear and nonlinear mixed effects models. R package version 3.1–143. https://CRAN.R-project.org/package=nlme
Pouteau R, van Kleunen M, Strasberg D (2023) Closely related aliens lead to greater extinction risk. Biol Conserv 284:110148
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Reinhart KO, Rinella MJ (2016) A common soil handling technique can generate incorrect estimates of soil biota effects on plants. New Phytol 210:786–789
Relva MA, Nuñez MA, Simberloff D (2010) Introduced deer reduce native plant cover and facilitate invasion of non-native tree species: evidence for invasional meltdown. Biol Invasions 12:303–311
Rinella MJ, Reinhart KO (2018) Toward more robust plant-soil feedback research. Ecology 99:550–556
Rutten G, Prati D, Hemp A, Fischer M (2016) Plant-soil feedback in East-African savanna trees. Ecology 97:294–301
Seebens H, Bacher S, Blackburn TM, Capinha C, Dawson W, Dullinger S, Genovesi P, Hulme PE, van Kleunen M, Kühn I, Jeschke JM, Lenzner B, Liebhold AM, Pattison Z, Pergl J, Pyšek P, Winter M, Essl F (2021) Projecting the continental accumulation of alien species through to 2050. Glob Change Biol 27:970–982
Seebens H, Blackburn TM, Dyer EE, Genovesi P, Hulme PE, Jeschke JM, Pagad S, Pyšek P, Winter M, Arianoutsou M, Bacher S, Blasius B, Brundu G, Capinha C, Celesti-Grapow L, Dawson W, Dullinger S, Fuentes N, Jager H, Kartesz J, Kenis M, Kreft H, Kuhn I, Lenzner B, Liebhold A, Mosena A, Moser D, Nishino M, Pearman D, Pergl J, Rabitsch W, Rojas-Sandoval J, Roques A, Rorke S, Rossinelli S, Roy HE, Scalera R, Schindler S, Štajerová K, Tokarska-Guzik B, van Kleunen M, Walker K, Weigelt P, Yamanaka T, Essl F (2017) No saturation in the accumulation of alien species worldwide. Nat Commun 8:14435
Simberloff D, Von Holle B (1999) Positive interactions of nonindigenous species: invasional meltdown? Biol Invasions 1:21–32
Thakur MP, van der Putten WH, Wilschut RA, Veen GF, Kardol P, van Ruijven J, Allan E, Roscher C, van Kleunen M, Bezemer TM (2021) Plant-Soil feedbacks and temporal dynamics of plant diversity-productivity relationships. Trends Ecol Evol 36:651–661
Van Grunsven RHA, Van Der Putten WH, Bezemer TM, Tamis WLM, Berendse F, Veenendaal EM (2007) Reduced plant-soil feedback of plant species expanding their range as compared to natives. J Ecol 95:1050–1057
van der Heijden MGA (2004) Arbuscular mycorrhizal fungi as support systems for seedling establishment in grassland. Ecol Lett 7:293–303
van der Heijden MGA, Bardgett RD, van Straalen NM (2008) The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett 11:296–310
van der Putten WH, Bardgett RD, Bever JD, Bezemer TM, Casper BB, Fukami T, Kardol P, Klironomos JN, Kulmatiski A, Schweitzer JA, Suding KN, Van de Voorde TFJ, Wardle DA, Hutchings M (2013) Plant-soil feedbacks: the past, the present and future challenges. J Ecol 101:265–276
van Kleunen M, Dawson W, Essl F, Pergl J, Winter M, Weber E, Kreft H, Weigelt P, Kartesz J, Nishino M, Antonova LA, Barcelona JF, Cabezas FJ, Cardenas D, Cardenas-Toro J, Castano N, Chacon E, Chatelain C, Ebel AL, Figueiredo E, Fuentes N, Groom QJ, Henderson L, Inderjit, Kupriyanov A, Masciadri S, Meerman J, Morozova O, Moser D, Nickrent DL, Patzelt A, Pelser PB, Baptiste MP, Poopath M, Schulze M, Seebens H, Shu WS, Thomas J, Velayos M, Wieringa JJ, Pyšek P (2015) Global exchange and accumulation of non-native plants. Nature 525:100–103
Vilà M, Espinar JL, Hejda M, Hulme PE, Jarošík V, Maron JL, Pergl J, Schaffner U, Sun Y, Pyšek P (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol Lett 14:702–708
Vilà M, Weiner J (2004) Are invasive plant species better competitors than native plant species? - evidence from pair-wise experiments. Oikos 105:229–238
Wang YJ, Müller-Schärer H, van Kleunen M, Cai AM, Zhang P, Yan R, Dong BC, Yu FH (2017) Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytol 216:1072–1078
Weigelt A, Jolliffe P (2003) Indices of plant competition. J Ecol 91:707–720
Wolfe BE, Klironomos JN (2005) Breaking new ground: soil communities and exotic plant invasion. Bioscience 55:477–487
Yang Q, Weigelt P, Fristoe TS, Zhang ZJ, Kreft H, Stein A, Seebens H, Dawson W, Essl F, Konig C, Lenzner B, Pergl J, Pouteau R, Pyšek P, Winter M, Ebel AL, Fuentes N, Giehl ELH, Kartesz J, Krestov P, Kukk T, Nishino M, Kupriyanov A, Villasenor JL, Wieringa JJ, Zeddam A, Zykova E, van Kleunen M (2021) The global loss of floristic uniqueness. Nat Commun 12:7290
Zhang Z, Liu Y, Brunel C, van Kleunen M (2020a) Evidence for Elton’s diversity-invasibility hypothesis from belowground. Ecology 101:e03187
Zhang Z, Liu Y, Brunel C, van Kleunen M (2020b) Soil-microorganism-mediated invasional meltdown in plants. Nat Ecol Evol 4:1612–1621
Zhang Z, van Kleunen M (2019) Common alien plants are more competitive than rare natives but not than common natives. Ecol Lett 22:1378–1386
Zuur AF, Ieno EN, Walker N, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
We thank Otmar Ficht, Claudia Martin, Yanran Dai, Ruben Schmidt, Louisa Neubauer, Christian Menn, Samuel Kundrat and Heinz Vahlenkamp for technical assistance. DC thanks funding from the China Scholarship Council (201906760016) and support from the International Max Planck Research School for Quantitative Behaviour, Ecology and Evolution.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
DC conceived the idea. MvK and DC designed the experiment, DC conducted the experiment and analyzed the data. DC wrote the first draft of the manuscript with further inputs from MvK.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Responsible Editor: Gaowen Yang.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
ESM 1
(PDF 717 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, D., van Kleunen, M. Negative conspecific plant-soil feedback on alien plants co-growing with natives is partly mitigated by another alien. Plant Soil (2024). https://doi.org/10.1007/s11104-024-06704-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11104-024-06704-9