
Abstract
Purpose of Review
Cyanobacteria, commonly known as blue-green algae, are often seen as a problem. Their accumulation (bloom) in surface water can cause toxicity and aesthetic concerns. Efforts have been made in preventing and managing cyanobacterial blooms. By contrast, purposeful cultivation of cyanobacteria can create great opportunities in food, chemical and biofuel applications. This review summarises the current stage of research and the socio-economic impacts associated with both the problems and opportunities induced from the presence of cyanobacteria in surface water.
Recent Findings
Insightful knowledge of factors that trigger cyanobacterial blooms has allowed for the development of prevention and control strategies. Advanced technologies are utilised to detect, quantify and treat cyanobacterial biomass and cyanotoxins in a timely manner. Additionally, understanding of cyanobacterial biochemical properties enables their applications in food and health industry, agriculture and biofuel production. Researchers have been able to genetically modify several cyanobacterial strains to obtain a direct pathway for ethanol and hydrogen production.
Summary
Cyanobacterial blooms have been effectively addressed with advances technologies and cyanobacterial research. However, this review identified a knowledge gap regarding cyanotoxin synthesis and the relevant environmental triggers. This information is essential for developing measures to prevent cyanobacterial blooms. Additionally, this review affirms the promising opportunities that cyanobacteria offer in the food, cosmetics, pigments and agriculture. Biofuel production from cyanobacterial biomass presents an immense potential but is currently constrained by the cultivation process. Thus, future research should strive to achieve effective mass harvesting of cyanobacterial biomass and obtain a profound understanding of cyanotoxin production.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cyanobacteria are a type of photosynthetic green-looking or blueish bacteria [1]. They possess chlorophyll a and release oxygen as a product of photosynthesis. The green pigment chlorophyll a together with other accessory pigments often cause a masking effect on the cyan (blue-green) hue of cyanobacterial pigment phycocyanin [2]. Cyanobacteria possess the oldest known fossils, dated back to 3.5 billion years ago. They have a significant contribution to the evolution and ecological change throughout the Earth’s history [3].
Harmful cyanobacterial blooms are a global problem. Found in a range of water environments (freshwater, coastal and marine), they have bloom-forming capabilities which pose significant concerns to the community. Cyanobacterial blooms are a threat to the drinking water supply due to their potential toxicity and the release of taste and odour compounds (e.g. geosmin and 2-methyl-isoborneol). Cyanotoxins produced from several common cyanobacteria are extreme risks to public health. Human or wildlife exposure to cyanotoxins can lead to severe illness, including death [4,5,6]. Additionally, harmful cyanobacterial blooms incur significant damage to the economy by disrupting the tourism and agricultural industry. Millions of dollars are spent every year to manage and control the impacts of cyanobacterial blooms [7].
New tools and technologies have been developed for managing cyanobacteria in surface water. For example, smart satellite imaging technique allows for early detection of cyanobacterial blooms [8]. Data collected from this system enables the authorities to develop real-time cyanobacterial bloom alerts useful for the general public and water suppliers. Types of cyanobacteria species and cyanotoxins can then be identified and possibly quantified from several approaches such as genetic techniques (e.g. quantitative polymerase chain reaction), biochemical assays and liquid chromatography [9, 10].
Apart from those aforementioned problems, cyanobacteria and their biomass also present great opportunities for the production of sustainable and valuable commodities. Cyanobacteria (e.g. Spirulina) are rich in proteins, vitamins and bioactive compounds [11]. Some cyanobacteria and their products are thus suitable to be consumed as food or health supplement. Useful bioactive compounds extracted from cyanobacteria were also shown to have natural antioxidant and water retention properties [12]. These make them great replacements for synthetic compounds often used in cosmetic formulations. Some cyanobacteria are capable of nitrogen-fixing; thus, cyanobacterial biomass from these species can be used as biofertilisers and soil conditioners [13]. Cyanobacterial biomass has also been actively explored as a potential feedstock to produce biofuels. Several studies have reported the production of ethanol, isobutanol and clean hydrogen from engineered cyanobacterial strains [14,15,16].
Global warming is likely to exacerbate harmful cyanobacterial bloom in both intensity and frequency [2]. The result can be both a threat of severe consequences of harmful cyanobacterial blooms and an opportunity to utilise them for beneficial applications. Several independent studies have underlined the problems or benefits of cyanobacteria and corresponding management strategies [2, 17,18,19]. Each of them was able to deliver useful and novel insights on a particular aspect of the topic. However, a complete overview on cyanobacteria in surface water and its impacts is inadequate. This paper aims to provide a full perspective of the topic, highlighting the current problems and opportunities associated with cyanobacteria in surface water, as well as the technologies used for cyanobacterial detection, control and harvesting. The knowledge gaps regarding cyanobacterial properties, cyanotoxins production and harvesting methods are also delineated through reviewing recent publications. This information is useful for the effective management of cyanobacteria and converting cyanobacterial biomass into valuable products.
Cyanobacteria
Cyanobacterial Classification and Phenotype
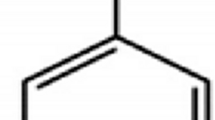
Cyanobacteria are photosynthetic prokaryotes which can grow in fresh, brackish and seawater [20]. There are about 2000 cyanobacterial species in 150 genera [21] such as Chroococcales, Anabaena and Nodularia. Cyanobacterial cells are identified to be more elaborated and larger than regular bacteria (0.5 to 60 μm in diameter). They are commonly found in unicellular, colonial and filamentous forms (Fig. 1) [22]. Their photosynthetic features and oxygen production set them apart from other bacteria. Several accessory pigments (e.g. chlorophyll a, phycocyanin and allophycocyanin) in the cells are light-harvesting antennae that capture the sunlight for energy in cyanobacterial photosynthesis.

The three common morphologies of cyanobacteria. Image courtesy of Landcare Research [23]
Most surface water cyanobacteria species have cells that contain gas vacuoles consisting of multiple gas vesicles. These gas vacuoles give cyanobacteria the buoyancy ability, i.e. the ability to remain in suspension and float to the water surface [24]. In highly stratified water or waters with great fluctuations in vertical mixing and optical depth, cyanobacteria with buoyancy-assisted vertical movements are favoured [25]. They are able to float to the water surface for optimal nutrients and light availability. Thus, their bloom forming capacity is enhanced and causes difficulty in removing cyanobacteria biomass from water (“Cyanobacterial Biomass Removal”).
Cyanobacteria possess CO2-concentrating mechanisms (CCMs) which consist of five inorganic carbon uptake systems. These enable the cells to increase the CO2 substrate for photosynthesis as well as the CO2 concentration in the cellular microcompartments for efficient enzyme operation [2]. The use of cyanobacteria for CO2 sequestration has been found to be very effective due to the presence of these CCMs [26, 27].
Another notable phenotype of many cyanobacteria is to fix atmospheric nitrogens (e.g. nitrogen-fixing cyanobacteria include Anabaena, Nostoc and Nodularia). Their cell structures (i.e. heterocyst) are thick wall which is impermeable to oxygen but permeable to nitrogen [28]. Nitrogen fixation is a competitive advantage for these filamentous cyanobacteria in a nitrogen-limited environment.
Cyanobacterial Bloom Triggers
Cyanobacterial blooms are the result of rapid and excessive growth as well as accumulation of cyanobacterial biomass on the water surface [2]. Because of the photosynthetic activity, environmental conditions that include nutrient availability, temperature, light exposure and CO2 are key factors to cyanobacterial growth [29, 30]. When these conditions are combined (e.g. often in summertime), cyanobacterial blooms will occur. Common harmful bloom-forming genera are Aphanizomenon, Cylindrospermopsis, Dolichospermum, Microcystis, Nodularia, Planktothrix and Trichodesmium [2].
Significant research efforts have been devoted to exploring the optimal growth conditions of cyanobacteria, which lead to blooms [31]. These conditions provide intuitive information to prevent and control cyanobacterial blooms (Sect. 4). For example, the eutrophic condition with low nitrogen to phosphorous ratio ranging from 10 to 15 is suggested to be optimal for cyanobacterial growth [22]. Literature data also indicates that phosphorus-rich water supports the dominance of cyanobacteria over other phytoplankton communities [32,33,34]. Smith (1983) hypothesised that by modifying the total nitrogen to total phosphorous ratio, cyanobacterial growth could be controlled [29]. This is due to the low count of cyanobacteria at nitrogen–phosphorous ratio greater than 29 to 1 by weight [29]. Light exposure (i.e. light intensity and duration) is another significant factor in determining the formation and duration of blooms [35]. Cyanobacteria contain a range of pigments such as chlorophyll a, allophycocyanin and phycocyanin which harvest light in the green, yellow and orange part of the spectrum. This range is much wider than that used by other phytoplankton species, giving cyanobacteria an advantage in terms of absorbing light for photosynthesis [22]. However, Montechiaro F. and Giordano M. (2006) had reported that some cyanobacteria (e.g. Phormidium autumnale) can hypernate without virtually any light for months and are able to thrive immediately following light exposure [36]. This emphasises the specific response and flexibility of individual cyanobacteria to light exposure. Water temperature (25 °C or above) is also favourable for cyanobacterial growth; thus, more severe blooms are observed in late spring throughout summer. In recent years, cyanobacterial blooms appear to occur earlier and last longer possibly as the result of climate change. Warm temperature reduces water viscosity, thus stimulating the sedimentation of competing larger, non-motile phytoplankton with weak floating ability [31]. O’Niel et al. [31] suggest that cyanobacteria have better competitiveness when stratification of water body occurs due to insular heating. The warmer upper water layer is more abundant in nutrients and light during stratification. Cyanobacteria can float upwards and utilise these factors for more rapid reproduction.
Rising atmospheric CO2 due to global warming also contributes to the severity of cyanobacterial blooms [2, 37]. It forms a steeper concentration gradient with the dissolved CO2, which has been depleted by cyanobacterial development. This leads to a greater influx of CO2 into the water body to reach equilibrium with the atmosphere [37]. Higher dissolved CO2 concentration intensifies bloom formation. Besides, the CCMs can utilise this availability of CO2 to enhance the function and growth of cyanobacteria.
Other factors influencing cyanobacterial blooms include water stratification and wind patterns [38, 39]. Stagnant water condition allows for a longer residence time of cyanobacterial cells. As a result, more nutrients, light radiation and CO2 are absorbed, increasing cyanobacterial growth rate. Light winds can expand the area of cyanobacterial scums [39] by driving them closer together and towards shores and bays. This increases the chances of human or animal in contact with the blooms which may contain harmful cyanotoxins. Awareness of such potential toxicity and measures for prevention and control of cyanobacterial blooms to be put in place are extremely important for the community.
Impact of Cyanobacterial Blooms
Cyanotoxin Production and Its Consequences
Cyanotoxins are secondary products (metabolites) from the metabolism of several cyanobacteria, most commonly Microcystis, Anabaena and Planktothrix genera [40,41,42]. They possess a variety of biological structures and induce a range of negative effects on human and animal health (Table 1). Cyanotoxins are usually produced and contained within the cyanobacterial cells (intracellular) [41]. Environmental stress or chemical and mechanical processes applied to cyanobacterial bloom can cause cell death, stimulating the release of cyanotoxins into the waterbody (extracellular). Benthic cyanobacteria such as Nostoc and Lyngbya do not occupy surface water but their occurrence and release of extracellular cyanotoxins into the surrounding water contribute to the harmful impacts of cyanobacterial blooms.
There are three main groups of cyanotoxins based on their chemical structure including cyclic peptides (e.g. microcystins and nodularins), alkaloids (e.g. neurotoxins and cylindrospermopsin) and lipopolysaccharides [4]. Cyanotoxins may also be classified into three groups according to their toxic effects: hepatotoxins, neurotoxins and dermototoxins (Table 1). Microcystins and nodularins are hepatotoxic cyclic peptides containing specific amino acids such as Adda (2S, 3S, 8S, 9S)-3-amino-9-methoxy-2,6,8-trimethyl1-10-pheny1deca-4,6-dienoic acid) [2]. Microcystins and nodularins inhibit the function of protein phosphatases, resulting in severe liver damage in human [2, 4]. Other health impacts could involve kidney and colon malfunctions, gastroenteritis, allergic and irritation reactions [42].
The alkaloid cyanotoxins include cylindrospermopsin, saxitoxins and anatoxins [4, 43, 44]. Cylindrospermopsin is a well-known hepatotoxin with the primary target of toxic effects being liver and kidney. Saxitoxins are representative neurotoxins of a large toxin family responsible for paralytic shellfish poisoning [45]. Saxitoxins are among the most potent toxins known. They can cause rapid paralysis by impairing the neuron systems and inhibiting muscle contraction [46]. Similarly, anatoxins and its variants anatoxin-a and anatoxin-a(s) are neurotoxins affecting the central nervous system. They can cause paralysis and asphyxiation (being oxygen-deprived) [40, 47]. Aplysiatoxins and lyngbyatoxin are representative of the dermatotoxin alkaloids. They can cause inflammation and severe dermatitis to people in contact with the filaments. These toxins are found in marine blue-green algae such as Planktothrix and Oscillatoria [48, 49]. They are potent tumour promoters and protein kinase C activators [50]. Lipopolysaccharides are the last groups of cyanotoxins classified by chemical structure. Lipopolysaccharides help to determine and maintain the shape and size of the cell [48]. Lipopolysaccharides trigger irritant and allergenic responses in mammals and tissues in contact with the toxins. They pose a significant concern for exposure due to their universal presence on the cell wall of a wide variety of cyanobacteria [4].
The distribution of cyanobacteria and cyanotoxins varies temporally and spatially. This could be due to the variations in the characteristics of cyanobacterial species and their preferred blooming conditions. Tropical Africa and Asia are abundant in bloom-forming genus Microcystis while Cylindrospermopsis is the most prevalent in Australia [17]. Both of these genera have occurred frequently in tropical America. As a result, there are differences in the cyanotoxins presented in these areas. The most common toxins in general are microcystis [41]. Cylindrospermopsin is the most frequently encountered cyanotoxin in Australia, and anatoxin is commonly found in Africa produced by blooms of Anabaena species [17].
Evidence on chronic health effects caused by exposure to recreational water infected with cyanobacterial bloom has been well documented in many parts of the world [4]. In the Paulo Afonso region of Brazil’s Bahia State, there were 2000 reported cases of gastroenteritis and 88 deaths over a period of 42 days in 1988. This was the result of Anabaena and Microcystis blooms in the newly constructed Itaparica Dam’s reservoir [6]. Lake Taihu, the third-largest freshwater lake in China supplying water for potable use, has also long been infected with harmful cyanobacterial blooms [5]. The presence of high concentration microcystin toxin from Microcystis spp. in untreated water (4.8–44.00 μg/L) in Lake Taihu has impaired the drinking water supplies. This concentration is higher than the upper limit of safe value for human exposure recommended by WHO (1 μg/L) (Table 1). Toxin residue was also detected in the treated tap water, which has been suggested to contribute to the prevalence of liver cancer in cities along Lake Taihu [51].
Concentration of cyanotoxins detected in a cyanobacterial bloom event is unpredictable and often exceeds the drinking water guidelines (Table 1). The guideline values represent the concentration at which the water is safe to drink over a lifetime consumption. For example, a provisional guideline value of 1.0 μg/L is recommended by WHO for microcystin-LR upper limit concentration in water [52]. Guideline calculation is based on the daily water intake, body weight and the concentration of toxins [52].
Water Quality
Cyanobacterial blooms affect and alter the characteristics and quality of the waterbody. These include the changes in the water colour, pH, dissolved oxygen (DO) level and the presence of unpleasant odours. In the long term, cyanobacterial blooms can have significant impacts on the ecosystems within the waterbody. The aesthetic value of recreational water bodies is reduced when cyanobacterial blooms occur. Accumulation of cyanobacterial scums on the water surface and along the shoreline is aesthetically displeasing. It can cause clear water to appear green and murky. Blooms can also result in earthy or musty odours and poor taste. This is caused by the production of taste and odour compounds (i.e. Geosmin and 2-methyl-isoborneol from cyanobacterial biomass). Anabaena, Planktothrix, Oscillatoria, Aphanizomenon, Lyngbya and Symploca are common species that contain known geosmin and 2-methyl-isoborneol [65, 66]. Human taste-and-odour detection threshold for these compounds are as low as 10 ng/L [67], making the presence of these in water for recreational use an unpleasant issue.
The water pH and DO level were also significantly affected by cyanobacterial blooms. During photosynthesis, cyanobacteria uptake carbon dioxide and consequently raise the pH by increasing the level of hydroxide. On the other hand, during the cyanobacteria cell lysis, pH level is reduced [68]. An increase or reduction in pH can be unfavourable for the ecosystems since many aquatic species prefer a stable pH range [69, 70]. Cyanobacteria uptake oxygen for their aerobic respiratory activities during the night time [71], causing the DO level to decrease. Degradation of dead cyanobacterial cells after blooms also requires oxygen [68]. The increase in biochemical oxygen demand (BOD) and oxygen depletion caused by cyanobacteria makes it more competitive for other aquatic species to thrive.
Socio-economic Impacts
Cyanobacterial blooms can disrupt the socio-economic stability due to their potential toxicity and impacts on water quality. Significant impacts are reported on industries such as tourism, agriculture, real estate and public health sector [72, 73]. Unfortunately, recent cost-analysis of these impacts is limited while available documents are dated back to the 1990s and 2000s. An example of a comprehensive report on financial damage caused by several cyanobacterial blooms was reviewed by Steffensen [7] for Australia in 2008 (Table 2). The impact on tourism was mainly due to the prohibition of recreational activities (e.g. fishing, camping, swimming) near bloom affected areas [7]. A study on cyanobacterial blooms from 1990 to 1999 in England and Wales [74] reported the damage costs to be $105–160 million per year. Dodds et al. estimated an annual economic loss of more than $1 billion in the USA due to harmful cyanobacterial blooms in 2008 [75]. Due to the rapid change in economic condition, the monetary values of socio-economic impacts in the event of cyanobacterial blooms are expected to be significantly higher for the recent years.
Monitoring measures to identify the presence and prevent the progression of cyanobacterial blooms also induce immediate costs. For example, Hamilton City Council in New Zealand spent $1000 a day in early 2003 to treat the city’s drinking water with powdered activated carbon in response to a potential saxitoxins bloom [76]. For cyanotoxin detection, toxicity tests may cost over $1000 per sample [7], although this cost will depend on the size of the blooms and facilities available for assessing.
The cost for the actions taken subsequently to control and remove the blooms (e.g. artificial mixing and algicides) is site-specific and could involve extra expenses. For example, $1 million is spent each year by South Australia Water to treat cyanobacterial blooms using copper-based algicide. This includes the dispose of the copper contaminated water treatment sludge as many aquatic organisms could be negatively impacted by copper [7]. Pretty et al. [74] also reported an expense of $77 million per year to address the damages from cyanobacterial blooms in England and Wales.
Cyanobacterial Bloom Control and Treatment
Effective cyanobacterial bloom control requires a holistic approach with well-integrated management and technology measures as well as a focus on the prevention strategies to minimise impact costs. Technologies are incorporated to effectively detect and control the development of cyanobacterial blooms and cyanotoxins (Fig. 2).

Cyanobacterial management response cycle
Prevention Strategies
The focus of preventive measures for cyanobacterial blooms includes (i) restricting the nutrient availability for cyanobacterial growth, (ii) facilitating changes in hydrodynamics that are unflavoured for blooms and (iii) making improvements towards climate change. Nutrient management tackles the root of the problem by limiting nutrient availability in the water bodies [2, 77]. Phosphorus has been identified as a key bloom promoting factor [78,79,80]. Recent studies have also highlighted the importance of nitrogen in supporting biomass and toxigenicity [81,82,83]. Measures to reduce external phosphorus inputs such as bans on phosphates in detergents, minimising the use of synthetic fertilisers and improved sewage treatment have been effective [2, 81]. In-lake methods such as hypolimnetic aeration and oxygenation to reduce internal phosphorus loading from sediments have succeeded in some cases [84,85,86]. In the 1990s, Australian CSIRO had developed lanthanum modified bentonite (commercially known as Phoslock®), an innovative phosphorous binding clay [87, 88]. Phoslock has proved to effectively remove total and soluble reactive phosphorus [89]. Meanwhile, due to its complex gaseous atmospheric cycle, nitrogen loading is more difficult to manage and can be costly [81, 90]. Constructions of wetlands and vegetative riparian buffers are effective ways to reduce non-diffuse sources of both phosphorus and nitrogen (e.g. agricultural and stormwater runoff) [91, 92]. These systems can also limit light availability to the surface cyanobacteria by shading the water and thus mitigate bloom development near shore.
Mixing can effectively mitigate cyanobacterial growth. Stagnant water allows buoyant cyanobacteria to remain stable in the upper layer abundant of light and warmth, thus promoting bloom development. By applying mixing, the stability of the water column is decreased while the mixing water depth is increased. As a result, cyanobacteria entrained in turbulence experience lower light availability, higher light fluctuation and shorter residence time. This leads to a decrease in cyanobacterial growth [18]. Some examples include introducing plumes of bubbles near the bottom of the reservoir or installing a propeller in/near the dam wall.
Sonication has emerged as a promising technique to control cyanobacterial bloom [93, 94]. Ultrasonic radiation in water can generate cavitation bubbles, causing localised regions of very high temperature and pressure [95]. This extreme environment disrupts the structure of the gas vacuoles in algal cells thus inhibiting the buoyancy of cyanobacteria. Without the competitive ability for buoyancy, the photosynthetic activity of cyanobacteria is limited. Simultaneously, the sedimentation of collapsed algal cells is stimulated [96]. This leads to a decline in cyanobacterial growth. Sonication is simple and easy to operate, with low impact on the ecosystems [93].
Improvements towards climate change and global warming also contribute to minimising cyanobacterial blooms in the long term. Currently, climate change affects cyanobacterial development in many ways [2, 81]. Higher water temperature due to global warming stimulates vertically thermal stratification, which favours the growth of cyanobacteria. Increased water variability (e.g. severe storms and rainfall) due to climate change may results in more nutrient runoff into the water bodies. Therefore, efforts in decelerating climate change can generate positive impacts on the long-term cyanobacterial bloom management. Successful prevention and management of cyanobacteria in surface water require the engagement of the wider community (e.g. water managers, users, scientists, engineers) [77]. Collaborations among these parties to develop a tailor-made and integrated solution for the water body of interest are encouraged.
Detection of Cyanobacterial Bloom Development
Remote sensing technologies (using satellite, drone and hyperspectral cameras) accompanied by advanced modelling (e.g. artificial neural networks) offers cost-effective and efficient ways to forecast and monitor cyanobacterial blooms [97, 98]. These have been implemented by CSIRO and Australian water authorities (WaterNSW and Melbourne Water) to develop a harmful algal bloom early warning system [8]. Based on historical data and the information collected from the hyperspectral camera, changes in the physical environment (temperature and wind) that could lead to cyanobacterial blooms are detected. This together with satellite remote sensing imagery allows for risk assessment of cyanobacterial blooms and their potential spatial spread [8]. Prediction of cyanobacterial development seven to 14 days ahead can be achieved through these approaches. This allows adequate time for early intervention (e.g. chemical dosing, apply mixing) to minimise a further growth of cyanobacteria [8].
Historical and current satellite data is also used to detect cyanobacterial blooms in US freshwater systems in a project called Cyanobacteria Assessment Network (CyAN). This project was facilitated in 2015 by multiple agencies, including the EPA, NASA, the National Oceanic and Atmospheric Administration (NOAA) and the United States Geological Survey (USGS) [99]. CyAN seeks to develop an integrated and reliable system to predict and identify cyanobacterial blooms across the US using satellites. The harmful level of these in various water storages can also be characterised using the data from colour satellites [99].
Public alerts for cyanobacteria detection provide up-to-date information for all water suppliers and users and prevent undesirable accidents related to cyanobacterial blooms. These could include media statements, signage and direct advice from the authorities. An online cyanobacteria alert system is particularly effective in delivering real-time update on the bloom development. The data collected from remote sensing is a good input for this type of systems.
An example of the online algal alert maps is provided by the Regional Algal Coordinating Committees (RACCs) across New South Wales, Australia [100]. Three colour-coded alert levels are used to represent the level of cyanobacteria in the water. They are declared once the algal cell numbers exceed the concentration in the Guidelines for Managing Risk in Recreational Waters [4] (Table 3).
Cyanobacterial Biomass Removal
A range of techniques has been used for removing and harvesting cyanobacterial biomass (and intracellular cyanotoxins) from a water suspension [101]. The most common techniques include coagulation and flocculation, flotation, membrane filtration and centrifugation (Table 4). The deployment of these techniques often focuses on several factors i.e. ease of use, removal efficiency, operational cost, energy demand, operation scale and quality of harvested algal biomass. It is also common to combine two or more techniques to achieve desirable operation and efficiency [102].
Cyanotoxin Detection and Removal
Identification of the toxicological potential of cyanobacterial blooms can be obtained through molecular techniques. These include polymerase chain reaction (PCR)–based methods and desoxyribonucleic Acid (DNA) microarrays [41, 107]. Conventional and real-time polymerase chain reaction (qPCR) methods are readily available and cost-effective ways to detect cyanotoxins, even at trace level [108]. Accurate detection is achieved through the amplification of the targeted toxin genes using primers if they are presented in the sample. The qPCR technique particularly allows for the indirect determination of the number of target genes [41, 108]. DNA microarrays are a recent technique used to detect and quantify cyanotoxins using their genomic DNA. Microcystins and nodularins have been identified using this technique [109, 110]. Although DNA microarray requires a high cost, it can provide rapid toxin detection [41].
Biochemical properties of cyanotoxins allow for the development of detection methods such as enzyme-linked immunosorbent assays (ELISA) and protein phosphatase inhibition assay (PPIA). Commercially available ELISA is an easy and inexpensive method for cyanotoxin testing [111]. ELISA relies on highly specific antibody-antigen interaction to detect cyanobacterial cells [112]. Currently, cyanotoxins that can be assessed and quantified by ELISA are microcystin, nodularin, cylindrospermopsin, saxitoxin and BMAA [112,113,114]. PPIA was developed based on the ability of cyanotoxins to inhibit the activity of protein phosphatase enzymes [41]. In the PPIA test, inhibition of protein phosphatase indicates toxin concentration. However, PPIA cannot distinguish among microcystin variants or between microcystin and nodularin despite them having a different reaction with phosphatase. The results, therefore, are expressed as equivalent MC-LR/L [41].
A range of chemical methods such as liquid-based separations and mass spectrometry is available for identification and quantification of cyanotoxins in water. Common cyanotoxins (e.g. Microcystins) can be identified using reversed-phase high-performance liquid chromatography (RP-HPLC). This allows for the separation of cyanotoxin molecules on the basis of hydrophobicity [115]. On the other hand, hydrophilic interaction liquid chromatography (HILIC) is useful for detecting very polar cyanotoxins (e.g. saxitoxins and β-N-methylamino-l-alanine (BMAA)) [41]. The mechanism involves the interaction between polar cyanotoxins and the stationary aqueous phase at the packing surface [116]. Recently, a combined RP-HPLC and HILIC system has emerged as a potential approach to simultaneously separate lipophilic and hydrophilic cyanotoxins [117]. Combination of liquid chromatography (e.g. HPLC or HILIC) and mass spectrometry (MS) is also a powerful analytical technique that delivers sensitive and selective results for toxin determination [112, 117]. These analytical methods provide accurate detection and quantification of cyanotoxins in environmental samples. However, a high level of expertise and expensive equipment is required to operate these systems [112].
Extracellular cyanotoxins can be removed from water using several techniques including ozonation, UV radiation and activated carbon (Table 5). These techniques are effective for degrading common cyanotoxins microcystis, cylindrospermopsin and anatoxin-a [40]. Cyanotoxin saxitoxins appear to be well removed using granular activated carbon [118]. Cyanobacterial blooms can contain several types of cyanotoxins; thus, it is recommended to combine these techniques for a better removal efficiency.
Green Liver Concept or Systems is a method to remove extracellular cyanotoxins based on plants’ capabilities to uptake, metabolise and store cyanotoxins in their cell wall fractions [119, 120]. This occurs during plants’ biotransformation process similar to that of animal’s liver. The suitable aquatic plants thus act as “green liver” to remediate contaminated water and pack away cyanotoxins at a low cost. Periodical harvesting of the aquatic macrophytes in this system is necessary to prevent the release of cyanotoxins from degrading plants back into the water [120]. Green Liver concept is a promising approach to sustainably remove extracellular cyanotoxins.
Multiple techniques (e.g. oxidation, coagulation, sedimentation and filtration) are often incorporated into a drinking water treatment design (Fig. 3) [102]. Different removal mechanisms offered by these techniques can assist one another, thus enhancing the total cyanobacteria and cyanotoxin removal efficiency. Pre-treatment techniques require careful consideration to avoid cell lysis and subsequent cyanotoxin release. For example, many drinking water treatments in Vietnam apply pre-chlorination as an algaecide. However, cyanobacterial cell’s membrane can be severely disrupted by chlorine, leading to cell lysis and extracellular cyanotoxins being liberated into drinking water [121]. The combination of treatment techniques can be determined by the quality of the water source. This data can be obtained through regular monitoring and testing across the plant. Long-term strategies to prevent cyanobacterial bloom in the source water should, therefore, be prioritised.

The schematic of drinking water treatment processes with considerations for effective cyanobacterial biomass and cyanotoxins removal based on recommendations and evaluations developed by EPA [102]
Opportunities from Cyanobacteria
The metabolic diversity allows cyanobacteria to possess a range of bioactive compounds. Not all of these bioactive compounds are toxic. Indeed, many of them are valuable for the food, health, cosmetic and pigment industry (Fig. 4). In addition, the photosynthetic capacity of cyanobacteria makes them one of the most promising feedstocks for solar-powered biofuel production (Fig. 4).

Potential applications of cyanobacterial biomass
Human Food
Cyanobacteria biomass has been a traditional food source for centuries. Kanembu people in central Africa and the indigenous population in Asia and North America are known to consume Spirulina as a nutritious food [19]. It is a high protein content (65%) superfood in comparison to most other natural food such as animal and fish flesh (15–25%), soybean (35%) and eggs (12%). Vitamins extracted from cyanobacterial biomass have also been used for health care. A rich amount of carotene, thiamine, riboflavin and vitamin B can be harvested from Spirulina [122].
Several secondary metabolites from cyanobacteria (polysaccharides, essential fat gamma-Linolenic acid) can be easily absorbed by human cell thus improving energy production. In addition, researchers have identified potential health-promoting effects from Spirulina consumption (e.g. suppression of hypertension and elevated serum glucose level, alleviation of hyperlipidaemia) [123, 124].
Commercial production of these cyanobacteria into “nutraceuticals” (food supplements marketed with nutritional and medicinal benefits) has been facilitated over that last few decades. Spirulina is a dominating species in commercial cyanobacterial biomass production with an estimated global output of 2000 t a year [125, 126]. The largest cultivation farm is in Hainan China (Hainan Simai Enterprising Ltd) and produces 200 t of Spirulina powder annually. Further research to optimise the harvesting and extraction of valuable compounds from cyanobacteria will establish an economic and environmental-friendly food industry for the future.
Essential Ingredients for Cosmetic Products
Bioactive compounds isolated from cyanobacteria are promising resources for natural cosmetic and skincare industry. For examples, mycosporine-like amino acids are potential compounds for the production of effective natural UV blockers due to their adsorption maxima in UV range [127, 128]. These are photoprotective compounds primarily engaged in the protection of cyanobacteria against detrimental solar radiation [129, 130]. Derivatives from mycosporine-like amino acids (e.g. tetrahydropyridines) as sunscreen pigments not only prevent damage from the UV radiation but also suppress inflammation and have antioxidant activity [131,132,133]. However, further research is needed to validate the industrial development of natural sunscreens and other cosmetic products from mycosporine-like amino acids.
Exopolysaccharides excreted from cyanobacteria (e.g. Synechocystis) have antioxidant properties and potential use as moisturising agents [134]. Exopolysaccharides are composed of various sugars and uronic acid with water adsorption and retention capacity [12]. In a comparison between a exopolysaccharides (sacran) extracted from Aphanothece sacrum and hyaluronic acid (the most widely used ingredient in moisturising products), Okajima et al. observed that sacran had a higher water absorption efficiency [135, 136]. Sacran, therefore, has the potential to replace expensive hyaluronic acid in the production of high moisturising products. Hence, the development of cosmetic formulations based on natural compounds from cyanobacteria is an ecologically friendly approach to provide skin benefits without inducing side effects and high cost like synthetic products [12].
Pigments as Natural Colourants
Major groups of light-harvesting pigments (chlorophyll, phycobiliproteins and carotenoids) produced by cyanobacteria are commercially valuable. Phycocyanin is a type of phycobiliproteins that have been used as natural colourants in food (e.g. chewing gum, ice cream, candies) [19]. It contains natural blue pigment and is abundant in cyanobacteria (e.g. Spirulina, Synechococcus, Anabaena) [11]. Phycocyanins as natural colourants are environment-friendly and eliminate potential health issues using synthetic colours (toxic, carcinogenic). Phycobilins (phycobiliproteins) are also used as chemical tags in research and in immunofluorescence technique. This is due to their ability to bind to specific antibodies and fluoresce at a particular wavelength [137].
Sustainable Agriculture
Cyanobacteria have promising applications in the field of sustainable agriculture due to their ability to fix atmospheric nitrogen (N2) in soil, enhance the solubility of nutrients and act as a soil conditioner [19, 138, 139]. A natural population of cyanobacteria is present in most paddy fields [138]. Rice fields with waterlogged conditions are especially favourable for their habitation [19, 138]. Cyanobacteria as biofertiliser in rice field can contribute to about 20–30 kg N/ha [140]. It thereby reduces the investment into chemical fertilisers without compromising with the normal yield. Nutrient availability (i.e. phosphorus) is also improved as cyanobacteria can solubilise and mobilise the insoluble organic phosphates present in the soil [13, 141]. The effect of cyanobacterial biofertiliser on crop growth is not spontaneous due to the gradual release of the fixed nitrogen into the soil. This enables the crops to utilise more nutrients available from the soil during growth stage [19, 138]. Examples of effective cyanobacterial biofertilisers include Anabaena variabilis, Nostoc muscorum, Aulosira fertissima and Tolypothrix tenuis [139].
The beneficial effects of cyanobacteria inoculation in crop field have also been reported for wheat, kale and willow [142,143,144]. These studies indicated that besides enhancing soil fertility, cyanobacteria are effective soil conditioners. Inoculation of cyanobacteria in sandy and calcareous soils improved the soil organic matter, water holding capacity and soil aggregate stability [142]. This was presumably due to the excretion of several compounds (polysaccharides, peptides, lipids, etc.) from cyanobacterial cells which helps in binding soil particles [19, 142]. Cyanobacteria are, therefore, economical options to replace expensive soil conditioners for common agricultural use.
Feedstocks for Biofuel Production
Several advantageous properties make cyanobacteria a good feedstock for solar-powered biofuel production [145]. Carbon rich biomass can be produced rapidly without competing for arable lands for food crops. Naturally available resources such as sunlight, water, atmospheric or water-dissolved CO2 are adequate for cyanobacterial growth. Some strains of cyanobacteria (e.g. Synechocystis sp., Synechococcus sp. and Anabaena sp.) can be easily and stably engineered for better biofuel production [145, 146]. Excretion of fuel outside the engineered cyanobacterial cells is also favoured over intracellular fuel production in eukaryotic algae [145].
Synthetic biology and metabolic engineering approaches have been introduced to cyanobacteria since they do not possess a complete biosynthetic pathway for biofuel production [147]. Deng and Coleman [148] transformed Synechococcus species with bacterial genes from Zymomonas mobilis to create a catalysed pathway for ethanol synthesis. These genes provided two key enzymes (pyruvate decarboxylase and alcohol dehydrogenase). They degraded sugars to pyruvate and then fermented it to produce ethanol and CO2 as the only products. The engineered Synechococcus sp. PCC 6803 yielded an ethanol concentration of 0.23 g/L [148]. A similar approach was applied by Gao et al. [14] to Synechococcus, with additional disruption to the biosynthetic pathway of poly-β-hydroxybutyrate. A significantly higher ethanol yield (5.50 g/L) was achieved by the transformed Synechococcus sp. PCC 6803 [14]. Another example of engineered cyanobacteria is the transgenic S. elongatus PCC 7942 for isobutanol production. An isobutanol yield of 0.45 g/L was achieved using this species via the artificial and non-fermentative pathway [15].
Cyanobacteria also produce molecular hydrogen (H2), a promising clean fuel for the future [145, 149]. The combustion of hydrogen for energy conversion does not result in any air pollution. Hydrogen has the highest energy per unit weight (141.65 MJ/kg) among all known fuels [145, 147, 150]. In cyanobacteria, nitrogenase enzymes have been reported to produce hydrogen most efficiently as a by-product of nitrogen fixation [16]. Several engineered cyanobacterial strains have been generated and evaluated for hydrogen production (Table 6).
Future Research Roadmap
The knowledge of cyanotoxin production is significant for solving problems regarding cyanobacterial bloom, but it is still not well understood. Cyanotoxin production is regulated by cyanobacterial gene and a variety of environmental factors such as nutrients, light, temperature and oxidants [155]. A few studies have been conducted to investigate the gene regulation and the impact of environmental factors on various toxin productions (e.g. microcystins, cylindrospermopsins, nodularins and saxitoxins) [156,157,158,159]. However, since these studies were conducted under different conditions and due to the complexity of each cyanobacterial system, it is difficult to establish a precise understanding of toxin synthesis and regulation [155]. Besides laboratory experiments, more field experiments should be conducted to determine the effect of environmental triggers on toxin production. Furthermore, advances in molecular research are required to obtain a clear view of toxin synthesis. Strong knowledge of toxin trigger and production will allow researchers to develop preventive measures or treatments against the presence of cyanotoxins in the water environment.
Cyanobacteria also present many opportunities. Some applications have been commercialised with demonstrated market value (e.g. Spirulina health food, biofertilisers and pigments) while the others are emerging as a potential (cosmetics and biofuel) (Fig. 5). Cyanobacteria biomass into biofuels can be an effective replacement to fossil fuels, but there are challenges in lowering the production cost. The cost of cyanobacterial cultivation and processing for biofuel production is still high, making biofuel from cyanobacterial biomass more expensive than fossil fuel. Besides, the use of genetically modify cyanobacteria species for biofuel production require careful considerations regarding potential environmental risks. Therefore, more research is still necessary to develop efficient and economically viable cultivation techniques and to gain further understanding of engineered cyanobacteria.

The current state of research on problems and opportunities associated with cyanobacteria in surface water
Conclusion
Cyanobacteria present significant threats to human health and the environment at the time of worsening climate change outlook. Harmful cyanobacterial blooms negatively cause impacts on the water quality, public health and the economy. Technologies have been effectively utilised to detect and remove cyanobacterial biomass and cyanotoxins from water. The key to mitigating cyanobacterial bloom and its consequences relies upon prevention strategies. It requires efforts in managing water quality and reducing global warming. Progress in the research area of cyanotoxin production will be useful for toxic bloom prevention. Despite the problems, there are also several and very significant opportunities from purposeful cyanobacteria cultivation and utilisation (e.g. agriculture, food, cosmetics and pigments). Recently, genetically engineered cyanobacteria attract attention as promising feedstocks for solar-powered biofuel production. Further research is still necessary to evaluate the application of these engineered cyanobacterial strains. Successful large-scale production of biofuels from cyanobacterial biomass will contribute significantly towards the global goal of sustainability.
References
Whitton BA, Potts M. Introduction to the cyanobacteria. In: Whitton BA, editor. Ecology of cyanobacteria II: their diversity in space and time. Dordrecht: Springer Netherlands; 2012. p. 1–13.
Huisman J, Codd GA, Paerl HW, Ibelings BW, Verspagen JMH, Visser PM. Cyanobacterial blooms. Nat Rev Microbiol. 2018;16(8):471–83. https://doi.org/10.1038/s41579-018-0040-1.
Introduction to the cyanobacteria University of California Museum of Paleontology. https://ucmp.berkeley.edu/bacteria/cyanointro.html. Accessed 11/11 2019.
NHMRC. Guidelines for managing risk in recreational waters Canberra: National Health and Medical Research Council; 2008.
Qin B, Li W, Zhu G, Zhang Y, Wu T, Gao G. Cyanobacterial bloom management through integrated monitoring and forecasting in large shallow eutrophic Lake Taihu (China). J Hazard Mater. 2015;287:356–63. https://doi.org/10.1016/j.jhazmat.2015.01.047.
Teixeira M, Costa M, Carvalho V, Pereira M, Hage E. Gastroenteritis epidemic in the area of the Itaparica Dam, Bahia, Brazil. Bull Pan Am Health Organ. 1993;27:244–53.
Steffensen DA. Economic cost of cyanobacterial blooms. In: Hudnell HK, editor. Cyanobacterial harmful algal blooms: state of the science and research needs. New York: Springer New York; 2008. p. 855–65.
CSIRO. Blue-green algae research 2019. https://www.csiro.au/en/Research/Environment/Water/Blue-green-algae/Our-research.
Zervou S-K, Christophoridis C, Kaloudis T, Triantis TM, Hiskia A. New SPE-LC-MS/MS method for simultaneous determination of multi-class cyanobacterial and algal toxins. J Hazard Mater. 2017;323:56–66. https://doi.org/10.1016/j.jhazmat.2016.07.020.
Sendall B, Reardon K, Hunt L, Menjivar T, Muhid P. Identification, detection & characterisation of cyanobacteria using traditional & DNA-based methods: forensic and scientific services, Queensland Government. 2019.
Udayan A, Arumugam M, Pandey A. Chapter 4 - nutraceuticals from algae and cyanobacteria. In: Rastogi RP, Madamwar D, Pandey A, editors. Algal green chemistry. Amsterdam: Elsevier; 2017. p. 65–89.
Morone J, Alfeus A, Vasconcelos V, Martins R. Revealing the potential of cyanobacteria in cosmetics and cosmeceuticals — a new bioactive approach. Algal Res. 2019;41:101541. https://doi.org/10.1016/j.algal.2019.101541.
Pathak J, Rajneesh MPK, Singh SP, Häder D-P, Sinha RP. Cyanobacterial farming for environment friendly sustainable agriculture practices: innovations and perspectives. Front Environ Sci. 2018;6(7). https://doi.org/10.3389/fenvs.2018.00007.
Gao Z, Zhao H, Li Z, Tan X, Lu X. Photosynthetic production of ethanol from carbon dioxide in genetically engineered cyanobacteria. Energy Environ Sci. 2012;5(12):9857–65. https://doi.org/10.1039/C2EE22675H.
Atsumi S, Hanai T, Liao JC. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature. 2008;451(7174):86–9. https://doi.org/10.1038/nature06450.
Allahverdiyeva Y, Leino H, Saari L, Fewer DP, Shunmugam S, Sivonen K, et al. Screening for biohydrogen production by cyanobacteria isolated from the Baltic Sea and Finnish lakes. Int J Hydrog Energy. 2010;35(3):1117–27. https://doi.org/10.1016/j.ijhydene.2009.12.030.
Mowe MAD, Mitrovic SM, Lim RP, Furey A, Yeo DCJ. Tropical cyanobacterial blooms: a review of prevalence, problem taxa, toxins and influencing environmental factors. J Limnol. 2015;74:205–24. https://doi.org/10.4081/jlimnol.2014.1005.
Visser PM, Ibelings BW, Bormans M, Huisman JJAE. Artificial mixing to control cyanobacterial blooms: a review. Aquat Ecol. 2016;50(3):423–41. https://doi.org/10.1007/s10452-015-9537-0.
Chakdar H, Jadhav S, Dhar D, Pabbi S. Potential applications of blue green algae. J Sci Ind Res. 2012;71:13–20. http://hdl.handle.net/123456789/13322.
Markou G, Georgakakis D. Cultivation of filamentous cyanobacteria (blue-green algae) in agro-industrial wastes and wastewaters: a review. Appl Energy. 2011;88(10):3389–401. https://doi.org/10.1016/j.apenergy.2010.12.042.
Vincent WF. Cyanobacteria. In: Likens GE, editor. Encyclopedia of inland waters. Oxford: Academic Press; 2009. p. 226–32.
Mur LR, Skulberg OM, Utkilen H. Cyanobacteria in the environment. In: Chorus I, Bartram J, editors. Toxic cyanobacteria in water: a guide to their public health consequences, monitoring and management: World Health Organization; 1999.
Cyanobacteria. Landcare Research https://www.landcareresearch.co.nz/resources/identification/algae/identification-guide/identify/guide/descriptions/cyanobacteria.
Manisha M. Cyanobacteria: occurrence, morphology and cell structure. http://www.biologydiscussion.com/bacteria/cyanobacteria/cyanobacteria-occurrence-morphology-and-cell-structure/52036.
Reynolds CS, Oliver RL, Walsby AE. Cyanobacterial dominance: the role of buoyancy regulation in dynamic lake environments. New Zeal J Mar Fresh. 1987;21(3):379–90. https://doi.org/10.1080/00288330.1987.9516234.
Kumar K, Dasgupta CN, Nayak B, Lindblad P, Das D. Development of suitable photobioreactors for CO2 sequestration addressing global warming using green algae and cyanobacteria. Bioresour Technol. 2011;102(8):4945–53. https://doi.org/10.1016/j.biortech.2011.01.054.
Upendar G, Singh S, Chakrabarty J, Chandra Ghanta K, Dutta S, Dutta A. Sequestration of carbon dioxide and production of biomolecules using cyanobacteria. J Environ Manag. 2018;218:234–44. https://doi.org/10.1016/j.jenvman.2018.04.031.
Atia A, Saad A. Review on freshwater blue-green algae (cyanobacteria): occurrence, classification and toxicology. Biosci Biotechnol Res Asia. 2014;11:1319–25. https://doi.org/10.13005/bbra/1522.
Smith VH. Low nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. Science. 1983;221(4611):669–71. https://doi.org/10.1126/science.221.4611.669.
Dao TS, Nimptsch J, Wiegand C. Dynamics of cyanobacteria and cyanobacterial toxins and their correlation with environmental parameters in Tri An Reservoir, Vietnam. J Water Health. 2016;14(4):699–712. https://doi.org/10.2166/wh.2016.257.
O’Neil JM, Davis TW, Burford MA, Gobler CJ. The rise of harmful cyanobacteria blooms: the potential roles of eutrophication and climate change. Harmful Algae. 2012;14:313–34. https://doi.org/10.1016/j.hal.2011.10.027.
Paerl HW, Huisman J. Climate change: a catalyst for global expansion of harmful cyanobacterial blooms. Environ Microbiol Rep. 2009;1(1):27–37. https://doi.org/10.1111/j.1758-2229.2008.00004.x.
Watson SB, McCauley E, Downing JA. Patterns in phytoplankton taxonomic composition across temperate lakes of differing nutrient status. Limnol Oceanogr. 1997;42(3):487–95. https://doi.org/10.4319/lo.1997.42.3.0487.
Trimbee AM, Prepas EE. Evaluation of total phosphorus as a predictor of the relative biomass of blue-green algae with emphasis on Alberta lakes. Can J Fish Aquat Sci. 1987;44(7):1337–42. https://doi.org/10.1139/f87-158.
Merel S, Walker D, Chicana R, Snyder S, Baurès E, Thomas O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ Int. 2013;59:303–27. https://doi.org/10.1016/j.envint.2013.06.013.
Montechiaro F, Giordano M. Effect of prolonged dark incubation on pigments and photosynthesis of the cave-dwelling cyanobacterium Phormidium autumnale (Oscillatoriales, Cyanobacteria). Phycologia. 2006;45(6):704–10. https://doi.org/10.2216/06-15.1.
Visser PM, Verspagen JMH, Sandrini G, Stal LJ, Matthijs HCP, Davis TW, et al. How rising CO2 and global warming may stimulate harmful cyanobacterial blooms. Harmful Algae. 2016;54:145–59. https://doi.org/10.1016/j.hal.2015.12.006.
Paerl HW. Mitigating harmful cyanobacterial blooms in a human- and climatically-impacted world. Life (Basel). 2014;4(4):988–1012. https://doi.org/10.3390/life4040988.
Wu T, Qin B, Brookes JD, Shi K, Zhu G, Zhu M, et al. The influence of changes in wind patterns on the areal extension of surface cyanobacterial blooms in a large shallow lake in China. Sci Total Environ. 2015;518–519:24–30. https://doi.org/10.1016/j.scitotenv.2015.02.090.
EPA, editor. Cyanobacteria and cyanotoxins: information for drinking water systems. Washington, DC; 2014.
Sanseverino I, Conduto António DS, Loos R, Lettieri T. Cyanotoxins: methods and approaches for their analysis and detection. Publications Office of the European Union. 2017. doi:http://doi.org/10.2760/630736 (print) http://doi.org/10.2760/36186 (online).
Zanchett G, Oliveira-Filho EC. Cyanobacteria and cyanotoxins: from impacts on aquatic ecosystems and human health to anticarcinogenic effects. Toxins (Basel). 2013;5(10):1896–917. https://doi.org/10.3390/toxins5101896.
D’Anglada LV, Strong J. Drinking water health advisory for the cyanobacterial toxin cylindrospermopsin. In: EPA US, editor. Washington, DC 2015.
Rosen BH. Understanding cyanobacterial ecological strategies p. https://www.epa.gov/sites/production/files/2016-05/documents/webinar-understanding-cyanobacterial.pdf.
Negri AP, Jones GJ. Bioaccumulation of paralytic shellfish poisoning (PSP) toxins from the cyanobacterium Anabaena circinalis by the freshwater mussel Alathyria condola. Toxicon. 1995;33(5):667–78. https://doi.org/10.1016/0041-0101(94)00180-G.
Wiese M, D'Agostino PM, Mihali TK, Moffitt MC, Neilan BA. Neurotoxic alkaloids: saxitoxin and its analogs. Mar Drugs. 2010;8(7):2185–211. https://doi.org/10.3390/md8072185.
Otten TG, Paerl HW. Health effects of toxic cyanobacteria in U.S. drinking and recreational waters: our current understanding and proposed direction. Curr Environ Health Rep. 2015;2(1):75–84. https://doi.org/10.1007/s40572-014-0041-9.
Sivonen K, Jones G. Cyanobacterial toxins. In: Bartram J, editor. Toxic cyanobacteria in water: a guide to their public health consequences, monitoring and management: World Health Organization; 1999.
Rzymski P, Poniedziałek B. Review paper: dermatotoxins synthesized by blue-green algae (cyanobacteria). Adv Dermatol Allergol. 2012;29(1):47–50.
Fujiki H, Suganuma M, Suguri H, Yoshizawa S, Takagi K, Nakayasu M et al. New tumor promoters from marine natural products. Marine toxins. ACS Symposium Series, vol 418: American Chemical Society; 1990. p. 232–40.
Chen Y, Yu S, Yang JB. Microcystins in drinking water and cancer mortality in a city along Taihu Lake. China Oncol. 2002;12:485–8.
Guidelines for drinking-water quality: forth edition incorporating the first addendum. 4th ed: World Health Organization; 2017.
D’Anglada LV, Donohue JM, Strong J, Hawkins B. In: EPA US, editor. Health effects support document for the cyanobacterial toxin microcystins. Washington, DC; 2015.
NHMRC, NHMMC. Australian drinking water guidelines paper 6 national water quality management strategy. Canberra: National Health and Medical Research Council, National Resource Management Ministerial Council; 2011.
Mazur-Marzec H, Sutryk K, Kobos J, Hebel A, Hohlfeld N, Błaszczyk A, et al. Occurrence of cyanobacteria and cyanotoxin in the Southern Baltic Proper. Filamentous cyanobacteria versus single-celled picocyanobacteria. Hydrobiologia. 2013;701(1):235–52. https://doi.org/10.1007/s10750-012-1278-7.
Volume 1: coastal and fresh waters. In: Guidelines for safe recreational water environments: World Health Organization; 2003.
Rücker J, Stüken A, Nixdorf B, Fastner J, Chorus I, Wiedner C. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon-dominated temperate lakes. Toxicon. 2007;50(6):800–9. https://doi.org/10.1016/j.toxicon.2007.06.019.
Saker ML, Eaglesham GK. The accumulation of cylindrospermopsin from the cyanobacterium Cylindrospermopsis raciborskii in tissues of the Redclaw crayfish Cherax quadricarinatus. Toxicon. 1999;37(7):1065–77. https://doi.org/10.1016/S0041-0101(98)00240-2.
Humpage AR, Falconer IR. Oral toxicity of the cyanobacterial toxin cylindrospermopsin in male Swiss albino mice: determination of no observed adverse effect level for deriving a drinking water guideline value. Environ Toxicol. 2003;18(2):94–103. https://doi.org/10.1002/tox.10104.
Trainer VL, Hardy FJ. Integrative monitoring of marine and freshwater harmful algae in Washington State for public health protection. Toxins (Basel). 2015;7(4):1206–34. https://doi.org/10.3390/toxins7041206.
Salmaso N, Cerasino L, Boscaini A, Capelli C. Planktic Tychonema (Cyanobacteria) in the large lakes south of the Alps: phylogenetic assessment and toxigenic potential. FEMS Microbiol Ecol. 2016;92(10). https://doi.org/10.1093/femsec/fiw155.
Ibelings BW, Backer LC, Kardinaal WEA, Chorus I. Current approaches to cyanotoxin risk assessment and risk management around the globe. Harmful Algae. 2015;49:63–74. https://doi.org/10.1016/j.hal.2014.10.002.
Solter PF, Beasley VR. Chapter 38 - phycotoxins. In: Haschek WM, Rousseaux CG, Wallig MA, editors. Haschek and Rousseaux’s handbook of toxicologic pathology. 3rd ed. Boston: Academic Press; 2013. p. 1155–86.
Lajeunesse A, Segura PA, Gélinas M, Hudon C, Thomas K, Quilliam MA, et al. Detection and confirmation of saxitoxin analogues in freshwater benthic Lyngbya wollei algae collected in the St. Lawrence River (Canada) by liquid chromatography–tandem mass spectrometry. J Chromatogr A. 2012;1219:93–103. https://doi.org/10.1016/j.chroma.2011.10.092.
Jüttner F, Watson SB. Biochemical and ecological control of Geosmin and 2-Methylisoborneol in source waters. Appl Environ Microbiol. 2007;73(14):4395–406. https://doi.org/10.1128/AEM.02250-06.
Izaguirre G, Hwang CJ, Krasner SW, McGuire MJ. Geosmin and 2-Methylisoborneol from cyanobacteria in three water supply systems. Appl Environ Microbiol. 1982;43(3):708–14.
Young WF, Horth H, Crane R, Ogden T, Arnott M. Taste and odour threshold concentrations of potential potable water contaminants. Water Res. 1996;30(2):331–40. https://doi.org/10.1016/0043-1354(95)00173-5.
Richardson K. Harmful or exceptional phytoplankton blooms in the marine ecosystem. In: Blaxter JHS, Southward AJ, editors. Advances in marine biology: Academic Press; 1997. p. 301–85.
Kong P, Moorman GW, Lea-Cox JD, Ross DS, Richardson PA, Hong C. Zoosporic tolerance to pH stress and its implications for Phytophthora species in aquatic ecosystems. Appl Environ Microbiol. 2009;75(13):4307–14. https://doi.org/10.1128/AEM.00119-09.
Atli G, Canli EG, Eroglu A, Canli M. Characterization of antioxidant system parameters in four freshwater fish species. Ecotoxicol Environ Saf. 2016;126:30–7. https://doi.org/10.1016/j.ecoenv.2015.12.012.
Schmetterer G. Cyanobacterial respiration. In: Bryant DA, editor. The molecular biology of cyanobacteria. Dordrecht: Springer Netherlands; 1994. p. 409–35.
Sanseverino I, Conduto António DS, Pozzoli L, Dobricic S, Lettieri T. Algal bloom and its economic impact. 2016.
Lopez CB, Jewett EB, Dortch Q, Walton BT, Hudnell HK. Scientific assessment of freshwater harmful algal bloom. Washington, DC; 2008.
Pretty JN, Mason CF, Nedwell DB, Hine RE, Leaf S, Dils R. Environmental costs of freshwater eutrophication in England and Wales. Environ Sci Technol. 2003;37(2):201–8. https://doi.org/10.1021/es020793k.
Dodds WK, Bouska WW, Eitzmann JL, Pilger TJ, Pitts KL, Riley AJ, et al. Eutrophication of U.S. freshwaters: analysis of potential economic damages. Environ Sci Technol. 2009;43(1):12–9. https://doi.org/10.1021/es801217q.
Kouzminov A, Ruck J, Wood SA. New Zealand risk management approach for toxic cyanobacteria in drinking water. 2007;31(3):275–81. https://doi.org/10.1111/j.1467-842X.2007.00061.x.
Ibelings BW, Fastner J, Bormans M, Visser PMJAE. Cyanobacterial blooms. Ecology, prevention, mitigation and control: editorial to a CYANOCOST Special Issue. Aquat Ecol. 2016;50(3):327–31. https://doi.org/10.1007/s10452-016-9595-y.
Schindler DW. Eutrophication and recovery in experimental lakes: implications for lake management. Science. 1974;184(4139):897–9. https://doi.org/10.1126/science.184.4139.897.
Jeppesen E, Jensen JP, Søndergaard M. Response of phytoplankton, zooplankton, and fish to re-oligotrophication: an 11 year study of 23 Danish lakes. Aquat Ecosyst Health. 2002;5(1):31–43. https://doi.org/10.1080/14634980260199945.
Schindler DW, Hecky RE, Findlay DL, Stainton MP, Parker BR, Paterson MJ, et al. Eutrophication of lakes cannot be controlled by reducing nitrogen input: results of a 37-year whole-ecosystem experiment. Proc Natl Acad Sci. 2008;105(32):11254–8. https://doi.org/10.1073/pnas.0805108105.
Paerl HW, Hall NS, Calandrino ES. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci Total Environ. 2011;409(10):1739–45. https://doi.org/10.1016/j.scitotenv.2011.02.001.
Harke MJ, Gobler CJ. Global transcriptional responses of the toxic cyanobacterium, Microcystis aeruginosa, to nitrogen stress, phosphorus stress, and growth on organic matter. PLoS One. 2013;8(7):e69834. https://doi.org/10.1371/journal.pone.0069834.
Müller S, Mitrovic SM. Phytoplankton co-limitation by nitrogen and phosphorus in a shallow reservoir: progressing from the phosphorus limitation paradigm. Hydrobiologia. 2015;744(1):255–69. https://doi.org/10.1007/s10750-014-2082-3.
Moore BC, Cross BK, Beutel M, Dent S, Preece E, Swanson M. Newman Lake restoration: a case study part III. Hypolimnetic oxygenation. Lake Reservoir Manage. 2012;28(4):311–27. https://doi.org/10.1080/07438141.2012.738463.
Toffolon M, Ragazzi M, Righetti M, Teodoru CR, Tubino M, Defrancesco C, et al. Effects of artificial hypolimnetic oxygenation in a shallow lake. Part 1: phenomenological description and management. J Environ Manag. 2013;114:520–9. https://doi.org/10.1016/j.jenvman.2012.10.062.
Bormans M, Maršálek B, Jančula DJAE. Controlling internal phosphorus loading in lakes by physical methods to reduce cyanobacterial blooms: a review. 2016;50(3):407–22. https://doi.org/10.1007/s10452-015-9564-x.
Douglas GB, Adeney JA, Robb M. A novel technique for reducing bioavailable phosphorus in water and sediments. International Association Water Quality Conference on Diffuse Pollution1999. p. 517–523.
Robb M, Greenop B, Goss Z, Douglas G, Adeney J. Application of PhoslockTM, an innovative phosphorus binding clay, to two Western Australian waterways: preliminary findings. Hydrobiologia. 2003;494(1):237–43. https://doi.org/10.1023/a:1025478618611.
Copetti D, Finsterle K, Marziali L, Stefani F, Tartari G, Douglas G, et al. Eutrophication management in surface waters using lanthanum modified bentonite: a review. Water Res. 2016;97:162–74. https://doi.org/10.1016/j.watres.2015.11.056.
Stroom JM, Kardinaal WEA. How to combat cyanobacterial blooms: strategy toward preventive lake restoration and reactive control measures. Aquat Ecol. 2016;50(3):541–76. https://doi.org/10.1007/s10452-016-9593-0.
Bullerjahn GS, McKay RM, Davis TW, Baker DB, Boyer GL, D’Anglada LV, et al. Global solutions to regional problems: collecting global expertise to address the problem of harmful cyanobacterial blooms. A Lake Erie case study. Harmful Algae. 2016;54:223–38. https://doi.org/10.1016/j.hal.2016.01.003.
Tátrai I, Mátyás K, Korponai J, Paulovits G, Pomogyi P. The role of the Kis-Balaton water protection system in the control of water quality of Lake Balaton. Ecol Eng. 2000;16(1):73–8. https://doi.org/10.1016/S0925-8574(00)00091-4.
Park J, Church J, Son Y, Kim K-T, Lee WH. Recent advances in ultrasonic treatment: challenges and field applications for controlling harmful algal blooms (HABs). Ultrason Sonochem. 2017;38:326–34. https://doi.org/10.1016/j.ultsonch.2017.03.003.
Rajasekhar P, Fan L, Nguyen T, Roddick FA. A review of the use of sonication to control cyanobacterial blooms. Water Res. 2012;46(14):4319–29. https://doi.org/10.1016/j.watres.2012.05.054.
Suslick KS. Sonochemistry. Science. 1990;247(4949):1439–45. https://doi.org/10.1126/science.247.4949.1439.
Schneider OD, Weinrich LA, Brezinski S. Ultrasonic treatment of algae in a New Jersey reservoir. J Am Water Works Assoc. 2015;107(10):E533–E42. https://doi.org/10.5942/jawwa.2015.107.0149.
Mishra S, Stumpf RP, Schaeffer BA, Werdell PJ, Loftin KA, Meredith A. Measurement of cyanobacterial bloom magnitude using satellite remote sensing. Sci Rep. 2019;9(1):18310. https://doi.org/10.1038/s41598-019-54453-y.
Bui MH, Pham TL, Dao TS. Prediction of cyanobacterial blooms in the Dau Tieng Reservoir using an artificial neural network. Mar Freshw Res. 2017;68(11):2070–80. https://doi.org/10.1071/MF16327.
Schaeffer B, Loftin K, Stumpf R, Werdell J. Agencies collaborate, develop a cyanobacteria assessment network. Eos. 2015;96. https://doi.org/10.1029/2015EO038809.
Algae. WaterNSW. 2019. https://www.waternsw.com.au/water-quality/algae#stay.
Treatment options. In: Newcombe G, editor. International guidance manual for the management of toxic cyanobacteria. United Kingdom: Global Water Research Coalition; 2009.
Waters T, Dugan A, Lieberman R, Speth T, Carroll G. In: EPA, editor. Water treatment optimization for cyanotoxins. Washington, DC: In; 2016.
Singh G, Patidar SK. Microalgae harvesting techniques: a review. J Environ Manag. 2018;217:499–508. https://doi.org/10.1016/j.jenvman.2018.04.010.
Griffiths MJ, Dicks RG, Richardson C, Harrison ST. Advantages and challenges of microalgae as a source of oil for biodiesel. In: Stoytcheva M, Montero G, editors. Biodiesel - feedstocks and processing technologies. InTechOpen; 2011.
Barros AI, Gonçalves AL, Simões M, Pires JCM. Harvesting techniques applied to microalgae: a review. Renew Sust Energ Rev. 2015;41:1489–500. https://doi.org/10.1016/j.rser.2014.09.037.
Fasaei F, Bitter JH, Slegers PM, van Boxtel AJB. Techno-economic evaluation of microalgae harvesting and dewatering systems. Algal Res. 2018;31:347–62. https://doi.org/10.1016/j.algal.2017.11.038.
Zaffiro A, Rosenblum L, Wendelken SC. Method 546: determination of total microcystins and nodularins in drinking water and ambient water by enzyme-linked immunosorbent assay: U.S. EPA; 2015.
Gaget V, Lau M, Sendall B, Froscio S, Humpage AR. Cyanotoxins: which detection technique for an optimum risk assessment? Water Res. 2017;118:227–38. https://doi.org/10.1016/j.watres.2017.04.025.
Rudi K, Skulberg OM, Skulberg R, Jakobsen KS. Application of sequence-specific labeled 16S rRNA gene oligonucleotide probes for genetic profiling of cyanobacterial abundance and diversity by array hybridization. Appl Environ Microbiol. 2000;66(9):4004–11. https://doi.org/10.1128/aem.66.9.4004-4011.2000.
Rantala A, Rizzi E, Castiglioni B, De Bellis G, Sivonen K. Identification of hepatotoxin-producing cyanobacteria by DNA-chip. Environ Microbiol. 2008;10(3):653–64. https://doi.org/10.1111/j.1462-2920.2007.01488.x.
EPA. Detection methods for cyanotoxins. 2005. https://www.epa.gov/ground-water-and-drinking-water/detection-methods-cyanotoxins.
Moreira C, Ramos V, Azevedo J, Vasconcelos V. Methods to detect cyanobacteria and their toxins in the environment. Appl Microbiol Biotechnol. 2014;98(19):8073–82. https://doi.org/10.1007/s00253-014-5951-9.
Kamp L, Church JL, Carpino J, Faltin-Mara E, Rubio F. The effects of water sample treatment, preparation, and storage prior to cyanotoxin analysis for cylindrospermopsin, microcystin and saxitoxin. Chem Biol Interact. 2016;246:45–51. https://doi.org/10.1016/j.cbi.2015.12.016.
Gurbuz F, Metcalf JS, Codd GA, Karahan AG. Evaluation of enzyme-linked immunosorbent assays (ELISAs) for the determination of microcystins in cyanobacteria. Environ Forensic. 2012;13(2):105–9. https://doi.org/10.1080/15275922.2012.676596.
Aguilar M-I. Reversed-phase high-performance liquid chromatography. In: Aguilar M-I, editor. HPLC of peptides and proteins: methods and protocols. Totowa: Springer New York; 2004. p. 9–22.
Dell’Aversano C, Hess P, Quilliam MA. Hydrophilic interaction liquid chromatography–mass spectrometry for the analysis of paralytic shellfish poisoning (PSP) toxins. J Chromatogr A. 2005;1081(2):190–201. https://doi.org/10.1016/j.chroma.2005.05.056.
Chen J, Gao L, Li Z, Wang S, Li J, Cao W, et al. Simultaneous screening for lipophilic and hydrophilic toxins in marine harmful algae using a serially coupled reversed-phase and hydrophilic interaction liquid chromatography separation system with high-resolution mass spectrometry. Anal Chim Acta. 2016;914:117–26. https://doi.org/10.1016/j.aca.2016.01.062.
Newcombe G, Nicholson B. Treatment options for the saxitoxin class of cyanotoxins. Water Supply. 2002;2(5–6):271–5. https://doi.org/10.2166/ws.2002.0179.
Nimptsch J, Wiegand C, Pflugmacher S. Cyanobacterial toxin elimination via bioaccumulation of MC-LR in aquatic macrophytes: an application of the “Green Liver Concept”. Environ Sci Technol. 2008;42(22):8552–7. https://doi.org/10.1021/es8010404.
Pflugmacher S, Kühn S, Lee S, Choi J, Baik S, Kwon K, et al. Green Liver Systems® for water purification: using the phytoremediation potential of aquatic macrophytes for the removal of different cyanobacterial toxins from water. Am J Plant Sci. 2015;6:1607–18. https://doi.org/10.4236/ajps.2015.69161.
Trung B, Dao TS, Faassen E, Lürling M. Cyanobacterial blooms and microcystins in Southern Vietnam. Toxins (Basel). 2018;10(11):471. https://doi.org/10.3390/2Ftoxins10110471.
Dillon JC, Phuc AP, Dubacq JP. Nutritional value of the alga Spirulina. In: Simopoulos AP, editor. Plants in human nutrition. Basel: Karger; 1995. p. 32–46.
Liang S, Liu X, Chen F, Chen Z. Current microalgal health food R & D activities in China. In: Ang PO, editor. Asian Pacific phycology in the 21st century: prospects and challenges. Developments in hydrobiology. Dordrecht: Springer Netherlands; 2004. p. 45–8.
Yamaguchi K. Recent advances in microalgal bioscience in Japan, with special reference to utilization of biomass and metabolites: a review. J Appl Phycol. 1996;8(6):487–502. https://doi.org/10.1007/bf02186327.
Spolaore P, Joannis-Cassan C, Duran E, Isambert A. Commercial applications of microalgae. J Biosci Bioeng. 2006;101(2):87–96. https://doi.org/10.1263/jbb.101.87.
Milledge JJ. Commercial application of microalgae other than as biofuels: a brief review. Rev Environ Sci Biotechnol. 2011;10(1):31–41. https://doi.org/10.1007/s11157-010-9214-7.
Oren A, Gunde-Cimerman N. Mycosporines and mycosporine-like amino acids: UV protectants or multipurpose secondary metabolites? FEMS Microbiol Lett. 2007;269(1):1–10. https://doi.org/10.1111/j.1574-6968.2007.00650.x.
Kageyama H, Waditee-Sirisattha R. Chapter 5 - mycosporine-like amino acids as multifunctional secondary metabolites in cyanobacteria: from biochemical to application aspects. In: Atta ur R, editor. Studies in natural products chemistry: Elsevier; 2018. p. 153–94.
Singh R, Parihar P, Singh M, Bajguz A, Kumar J, Singh S, et al. Uncovering potential applications of cyanobacteria and algal metabolites in biology, agriculture and medicine: current status and future prospects. Front Microbiol. 2017;8(515). https://doi.org/10.3389/fmicb.2017.00515.
Liu Z, Häder DP, Sommaruga R. Occurrence of mycosporine-like amino acids (MAAs) in the bloom-forming cyanobacterium Microcystis aeruginosa. J Plankton Res. 2004;26(8):963–6. https://doi.org/10.1093/plankt/fbh083.
Bhatia S, Garg A, Sharma K, Kumar S, Sharma A, Purohit AP. Mycosporine and mycosporine-like amino acids: a paramount tool against ultra violet irradiation. Pharmacogn Rev. 2011;5(10):138–46. https://doi.org/10.4103/0973-7847.91107.
Stevenson CS, Capper EA, Roshak AK, Marquez B, Grace K, Gerwick WH, et al. Scytonemin-a marine natural product inhibitor of kinases key in hyperproliferative inflammatory diseases. Inflamm Res. 2002;51(2):112–4. https://doi.org/10.1007/bf02684014.
Rastogi RP, Sonani RR, Madamwar D. Cyanobacterial sunscreen Scytonemin: role in photoprotection and biomedical research. Appl Biochem Biotechnol. 2015;176(6):1551–63. https://doi.org/10.1007/s12010-015-1676-1.
De Philippis R, Sili C, Paperi R, Vincenzini M. Exopolysaccharide-producing cyanobacteria and their possible exploitation: a review. J Appl Phycol. 2001;13(4):293–9. https://doi.org/10.1023/A:1017590425924.
Okajima MK, Bamba T, Kaneso Y, Hirata K, Fukusaki E, Kajiyama S, et al. Supergiant ampholytic sugar chains with imbalanced charge ratio form saline ultra-absorbent hydrogels. Macromolecules. 2008;41(12):4061–4. https://doi.org/10.1021/ma800307w.
Okajima MK, Miyazato S, Kaneko T. Cyanobacterial megamolecule sacran efficiently forms LC gels with very heavy metal ions. Langmuir. 2009;25(15):8526–31. https://doi.org/10.1021/la8036956.
Saini DK, Pabbi S, Shukla P. Cyanobacterial pigments: perspectives and biotechnological approaches. Food Chem Toxicol. 2018;120:616–24. https://doi.org/10.1016/j.fct.2018.08.002.
Pabbi S. Blue green algae: a potential biofertilizer for rice. In: Sahoo D, Seckbach J, editors. The algae world. Dordrecht: Springer Netherlands; 2015. p. 449–65.
Singh JS, Kumar A, Rai AN, Singh DP. Cyanobacteria: a precious bio-resource in agriculture, ecosystem, and environmental sustainability. 2016;7(529). https://doi.org/10.3389/fmicb.2016.00529.
Issa A, Abd-Alla M, Ohyama T. Nitrogen fixing cyanobacteria: future prospect. In: Ohyama T, editor. Advances in biology and ecology of nitrogen fixation. InTechOpen; 2014. p. 23–48.
Rai AK, Sharma NK. Phosphate metabolism in the cyanobacterium Anabaena doliolum under salt stress. Curr Microbiol. 2006;52(1):6–12. https://doi.org/10.1007/s00284-005-0043-9.
Ghazal FM, Mahdy E-SM, MSA EL-F, EL-Sadany AEGY, Doha NME. The use of cyanobacteria as biofertilizer in wheat cultivation under different nitrogen rates. Nat Sci. 2018;16(4):30–5. https://doi.org/10.7537/marsnsj160418.06.
Gebre E, Abdi T, Wolde-meskel E, Bulta A, Davis J. Response of kale (Brassica oleracea L) crop to cyanobacterial biofertilizer in Ziway area, Ethiopia. Journal of Biology, Agriculture and Healthcare. 2018;8(13).
Grzesik M, Romanowska-Duda Z, Kalaji HM. Effectiveness of cyanobacteria and green algae in enhancing the photosynthetic performance and growth of willow (Salix viminalis L.) plants under limited synthetic fertilizers application. Photosynthetica. 2017;55(3):510–21. https://doi.org/10.1007/s11099-017-0716-1.
Sarsekeyeva F, Zayadan BK, Usserbaeva A, Bedbenov VS, Sinetova MA, Los DA. Cyanofuels: biofuels from cyanobacteria. Reality and perspectives. Photosynth Res. 2015;125(1):329–40. https://doi.org/10.1007/s11120-015-0103-3.
Savakis P, Hellingwerf KJ. Engineering cyanobacteria for direct biofuel production from CO2. Curr Opin Biotechnol. 2015;33:8–14. https://doi.org/10.1016/j.copbio.2014.09.007.
Singh V, Chaudhary DK, Mani I, Dhar PK. Recent advances and challenges of the use of cyanobacteria towards the production of biofuels. Renew Sust Energ Rev. 2016;60:1–10. https://doi.org/10.1016/j.rser.2016.01.099.
Deng M-D, Coleman JR. Ethanol synthesis by genetic engineering in cyanobacteria. Appl Environ Microbiol. 1999;65(2):523–8.
Sveshnikov DA, Sveshnikova NV, Rao KK, Hall DO. Hydrogen metabolism of mutant forms of Anabaena variabilis in continuous cultures and under nutritional stress. FEMS Microbiol Lett. 1997;147(2):297–301. https://doi.org/10.1111/j.1574-6968.1997.tb10257.x.
Ali I, Basit MA. Significance of hydrogen content in fuel combustion. Int J Hydrog Energy. 1993;18(12):1009–11. https://doi.org/10.1016/0360-3199(93)90083-M.
Srirangan K, Pyne ME, Perry CC. Biochemical and genetic engineering strategies to enhance hydrogen production in photosynthetic algae and cyanobacteria. Bioresour Technol. 2011;102(18):8589–604. https://doi.org/10.1016/j.biortech.2011.03.087.
Cournac L, Guedeney G, Peltier G, Vignais PM. Sustained photoevolution of molecular hydrogen in a mutant of Synechocystis sp. strain PCC 6803 deficient in the type I NADPH-dehydrogenase complex. J Bacteriol. 2004;186(6):1737. https://doi.org/10.1128/JB.186.6.1737-1746.2003.
Yoshino F, Ikeda H, Masukawa H, Sakurai H. High photobiological hydrogen production activity of a Nostoc sp. PCC 7422 uptake hydrogenase-deficient mutant with high nitrogenase activity. Mar Biotechnol. 2007;9(1):101–12. https://doi.org/10.1007/s10126-006-6035-3.
Mona S, Kaushik A, Kaushik CP. Hydrogen production and metal-dye bioremoval by a Nostoc linckia strain isolated from textile mill oxidation pond. Bioresour Technol. 2011;102(3):3200–5. https://doi.org/10.1016/j.biortech.2010.11.005.
Boopathi T, Ki J-S. Impact of environmental factors on the regulation of cyanotoxin production. Toxins (Basel). 2014;6(7):1951–78. https://doi.org/10.3390/toxins6071951.
Alexova R, Haynes PA, Ferrari BC, Neilan BA. Comparative protein expression in different strains of the bloom-forming cyanobacterium Microcystis aeruginosa. Mol Cell Proteomics. 2011;10(9):M110.003749. https://doi.org/10.1074/mcp.M110.003749.
Camargo S, Valladares A, Flores E, Herrero A. Transcription activation by NtcA in the absence of consensus NtcA-binding sites in an anabaena heterocyst differentiation gene promoter. J Bacteriol. 2012;194(11):2939–48. https://doi.org/10.1128/JB.05994-11.
Burford MA, Davis TW, Orr PT, Sinha R, Willis A, Neilan BA. Nutrient-related changes in the toxicity of field blooms of the cyanobacterium. Cylindrospermopsis raciborskii. 2014;89(1):135–48. https://doi.org/10.1111/1574-6941.12341.
Tsuchiya S, Cho Y, Konoki K, Nagasawa K, Oshima Y, Yotsu-Yamashita M. Synthesis and identification of proposed biosynthetic intermediates of saxitoxin in the cyanobacterium Anabaena circinalis (TA04) and the dinoflagellate Alexandrium tamarense (Axat-2). Org Biomol Chem. 2014;12(19):3016–20. https://doi.org/10.1039/C4OB00071D.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Water Pollution
Rights and permissions
About this article

Cite this article
Vu, H.P., Nguyen, L.N., Zdarta, J. et al. Blue-Green Algae in Surface Water: Problems and Opportunities. Curr Pollution Rep 6, 105–122 (2020). https://doi.org/10.1007/s40726-020-00140-w
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40726-020-00140-w