
3.2: Proto-homínidos
- Última actualización
- Guardar como PDF
- Page ID
- 139346

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Al determinar qué características fósiles denotan que un espécimen es un homínido, se examinan muchas características diferentes, incluidas las relacionadas con el bipedalismo, del que aprendiste anteriormente, y la masticación no afilada. Los simios tienen un complejo de bruñido canino-premolar, lo que significa que existe un diastema entre el tercer premolar inferior canino inferior donde encaja el canino superior cuando se cierran las mandíbulas (Larsen 2014). También se escrutan otras características como el tamaño cerebral y corporal y qué comportamientos culturales, en su caso, están asociados con los restos fósiles.
Las diversas características asociadas a los homínidos se desarrollaron a diferentes ritmos, situación que se conoce como evolución del mosaico. Para determinar a qué especie pertenece un espécimen fósil, los paleoantropólogos utilizan anatomía comparada. Hay mucha discusión entre los investigadores sobre el número exacto de especies. Algunos tienden a ver amplios rangos de variación, lo que resulta en menos especies. Los investigadores con este enfoque a veces se denominan “terradores” porque abultan especímenes fósiles en relativamente pocas especies. Otros investigadores ven rangos de variación más estrechos y se les conoce como “divisores” porque ven más variación de especies en el registro fósil. Para los fines de este curso, vamos a utilizar el enfoque presentado en Jurmain et al. (2013), que es más un enfoque de lumper.
Si bien los homínidos se presentarán más o menos en orden cronológico, no confundan el orden cronológico con las relaciones evolutivas lineales; algunos homínidos que se presentan no están en la línea directa con los humanos modernos. También es importante tener en cuenta que cada año se hacen nuevos descubrimientos que refinan lo que sabemos sobre la evolución humana. Lo que esto significa es que lo que aprendas este trimestre podría cambiar algo el próximo año a medida que se pongan a disposición nuevos datos. Si bien esto puede hacer que estudiar la evolución humana sea frustrante a veces, los nuevos descubrimientos nos ayudan a comprender mejor cómo evolucionaron nuestros primeros antepasados.
Es muy recomendable que comiences tu exploración de la evolución humana viendo el documental, Becoming Human.
Proto-homínidos
Sahelanthropus tchadensis
Descubierto por la Misión Paléoantropológica Franco-Tchadienne (MPFT) dirigida por Michel Brunet en Toros-Menalla, Chad, durante la temporada de campo 2001-2002, Sahelanthropus tchadensis es el homínido más antiguo descubierto hasta la fecha. La datación de nuclidos cosmogénicos (https://en.Wikipedia.org/wiki/Surface_exposure_dating) coloca a Sahelanthropus entre 7.2 a 6.8 mya (hace millones de años) (Lebatard et al. 2008). El binomio reconoce la zona donde se encontraron los fósiles, Sahel. El espécimen tipo, también denominado holotipo, incluye “... un cráneo casi completo y... partes de dos mandíbulas y algunos dientes aislados” (Bailey 2006:2044).
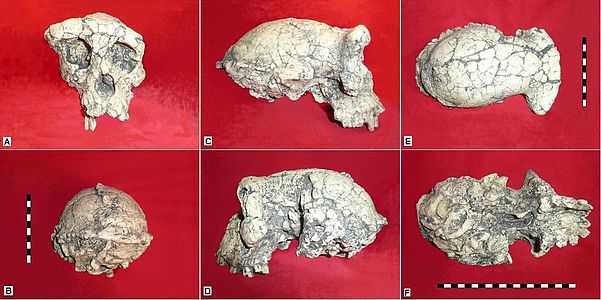
Figura\(\PageIndex{1}\) - Sahelanthropus tchadensis
El cráneo es una combinación de rasgos parecidos a simios y humanos. Las características similares a los simios incluyen el tamaño del cerebro, la cresta pesada de la frente Sus características similares a los humanos incluyen la posición delantera del foramen magnum (visite http://www.nature.com/scitable/knowl...hecus-67648286 (desplácese hacia abajo) para ver la imagen comparando la ubicación del foramen magnum en chimpancés, humanos y Sahelanthropus), prognatismo reducido, dientes caninos más pequeños y espesor intermedio del esmalte premolar y molar. Debido a la pronunciada cresta de la frente, el equipo de Brunet sugiere que el espécimen de Sahelanthropus es masculino.
Existe un debate entre los investigadores sobre si Sahelanthropus es un homínido o un simio. Algunos sugieren que el espécimen pertenece al de un simio hembra porque es probable que encuentre caninos usados en las puntas en simios hembra. Los críticos afirman que sin evidencia postcraneal es prematuro afirmar que Sahelanthropus es bípedo (Su 2013).
Los datos paleoecológicos sugieren que este espécimen de Sahelanthropus murió cerca de la orilla del lago y el área fue ampliamente boscosa.
Orrorin tugenensis
Encontrado en 2000 por un equipo liderado por Martin Pickford y Brigitte Senut, Orrorin tugenensis está representado por una colección de fósiles que incluyen un mínimo de cinco individuos. Se encontraron fragmentos de numerosas partes fosilizadas del cuerpo: mandíbula, fémur, húmero y huesos de dedos. Se encontraron muchos dientes, incluyendo molares y caninos. El binomio refleja dónde se encontraron los fósiles, la región de Tugen Hills en Kenia. Orrorin significa “hombre original” en el dialecto Tugen. Usando múltiples técnicas de datación, incluyendo paleomagnetismo y biocronología, los fósiles de Tugen Hills están fechados en aproximadamente 6.0 mya.
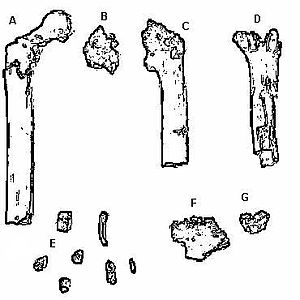
Figura\(\PageIndex{2}\) - Fósiles de Orrorin tugenensis
Al igual que Sahelanthropus, Orrorin tiene una mezcla de rasgos parecidos a simios y humanos. Sus dientes son más simios, por lo que la determinación de la bipedalidad es fundamental para atribuir los fósiles a la de un homínido. El equipo de investigación fundador afirma que las características del fémur indican que Orrorin era un bípedo y en la línea directa con los humanos modernos. Estudios más recientes sugieren que Orrorin era de hecho un homínido bípedo, pero no en la línea directa con Homo (Richmond y Jungers 2008).
La evidencia paleoecológica asociada a los fósiles de Orrorin sugiere que el área era un bosque poblado de lagos y arroyos.
Ardipithecus
Se han identificado dos especies diferentes de Ardipithecus: Ar. kaddaba y Ar. ramidus. Las dos especies se diferencian principalmente en el complejo C/P3 (características canino-premolares), específicamente, que los caninos de A r. kaddaba son más primitivos que los de Ar. ramidus.
Ar. kaddaba es la más antigua de las dos especies. Tiene fecha de 5.8-5.2 mya. Tim White, Gen Suwa y Yohannes Haile Selassie descubrieron Ar. fósiles de kaddaba en la región de Middle Awash de Etiopía. La colección fósil está compuesta por fragmentos de mandíbula y dientes, antebrazo, clavícula y cuarto dedo del pie. La forma del canino superior es más parecida a la humana que a los simios, ya que son más pequeños con una corona más circular. En general, los fragmentos postcraneales son más parecidos a simios; sin embargo, el hueso del dedo del pie mencionado anteriormente es similar a los huesos del dedo del pie de Australopithecus afarensis, una especie que tenía todas las señas de identidad del bipedalismo habitual. Otros investigadores afirman que un solo hueso del cuarto dedo del pie no proporciona evidencia suficiente para confirmar que Ar. kaddaba era un homínido bípedo.

Figura\(\PageIndex{3}\) - Ardipithecus ramidus: Esqueleto de Ardi
Los datos paleoecológicos sugieren que Ar. kaddaba vivía en un “hábitat cerrado y densamente boscoso cercano a fuentes permanentes de agua (por ejemplo, lagos y/o ríos) con condiciones pantanosas y pastizales de llanuras aluviales” (Becoming Human c2008).
A Tim White también se le atribuye el descubrimiento de Ar. ramidus en Aramis, Etiopía. Datado alrededor de 4.4 mya, Ar. ramidus “representa la especie más antigua que posee rasgos inequívocamente vinculados al linaje de los homínidos. Así, Ar. ramidus es la mejor evidencia descubierta hasta ahora para la raíz del árbol genealógico de los homínidos” (Becoming Human c2008). Los paleoantropólogos conocen más sobre Ar. ramidus que las especies mencionadas anteriormente, ya que 110 ejemplares han sido desenterrados en Aramis.
Postcranealmente, Ar. ramidus sigue siendo simio; sin embargo, hay algunas características que sugieren que la especie podría caminar bipedalmente, pero lo más probable es que pasara tiempo en los árboles al igual que los chimpancés modernos. En otras palabras, sus características de mosaico sugieren bipedalismo facultativo. Las características que llevan a los investigadores a afirmar que Ar. ramidus podría caminar bipedalmente incluyen la posición del foramen magnum y las características de la columna vertebral y el ilion.
Homínidos tempranos
Australopithecus anamensis
Australopithecus anamensis, que significa “simio sureño del lago”, fue encontrado por Maeve Leakey y Alan Walker en 1994 en Kanapoi, Kenia. Se recuperaron fósiles adicionales de A u. anamensis en la bahía de Allia, Kenia y varios sitios en Etiopía: Asa Issie, Woranso-Mille y Fejej. Los especímenes fósiles incluyen mandíbulas, maxilar superior, dientes, húmero parcial, tibia, falange manual, capitado, fémur parcial, varias vértebras y un hueso temporal parcial (Ward et al 2013). Usando métodos de datación radioisotópica en depósitos volcánicos, Au. anamensis data de 4.1-3.9 mya. La evidencia fósil sugiere que Au. anamensis es ancestral de Au. afarensis.

Figura\(\PageIndex{4}\) - Fósiles de Australopithecus anamensis
Au. anamensis exhibe un mosaico de rasgos parecidos a simios y homos. El tamaño y la forma de los dientes, especialmente el premolar inferior y el canino superior, la arcada dental en forma de U y el prognatismo son algunas de las características parecidas a simios, mientras que el grosor del esmalte y la amplitud de los molares se parecen más al Homo temprano. El único hueso de la muñeca también es como Homo. Los huesos de las extremidades inferiores sugieren que la anamensis Au. fue completamente bípeda.
Los datos paleoecológicos sugieren que Au. anamensis vivió en áreas boscosas cercanas a arroyos.
Australopithecus afarensis
Descubierto por Donald Johanson en 1974 en Hadar, Etiopía, Australopithecus afarensis es posiblemente la especie de homínidos fósiles más conocida. Está fechado de 3.7-3.0 mya (Scarre 2014). El descubrimiento de Johanson en 1974 fue, en su momento, el fósil de homínidos más completo. Se recuperó más del 40% del esqueleto, lo que permitió al equipo reconstruir completamente el esqueleto. Este espécimen fósil, llamado Lucy, aunado a las huellas encontradas en Laetoli, Tanzania, en 1978 por Mary Leakey, confirmó que Au. afarensis era completamente bípedo, aunque no exactamente como los humanos modernos. Las huellas en Laetoli indican que Au. afarensis tuvo un paso corto y una marcha paseante. Desde la década de 1970, se han encontrado cientos de especímenes de Au. afarensis (¡60 individuos al menos solo de Hadar!) en Etiopía, Kenia y Tanzania, permitiendo a los paleoantropólogos hacer “declaraciones definitivas [el énfasis original] sobre el patrón locomotor y la estatura de” (Jurmain 2013:211) este homínido temprano.

Figura\(\PageIndex{5}\) - Reconstrucción del esqueleto fósil de “Lucy” el Australopithecus afarensis
Au. afarensis tiene varias características primitivas o similares a simios, incluyendo un cerebro relativamente pequeño en comparación con Homo, una arcada dental en forma de U, una nariz plana, una frente aplanada (conocida como platicefalia) y una cara prognatica. Sus caninos, si bien son más grandes que el Homo, son más pequeños que los homínidos anteriores. Si bien su cerebro era más grande que los homínidos anteriores, todavía es pequeño en comparación con Homo. Hay evidencia de dimorfismo sexual; los machos Au. afarensis no eran más altos de 5 pies y las hembras alrededor de 3-4 pies, similares en proporción a los simios africanos modernos. Au. afarensis tiene un conjunto de rasgos derivados asociados con el bipedalismo; es decir, pelvis en forma de cuenco, curva en S de la columna vertebral y anatomía de la rodilla. Sin embargo, la curvatura de los dedos de las manos y los pies y la proporción de brazos a piernas sugieren a algunos investigadores que Au. afarensis pasó algún tiempo en los árboles.
En 2006, Zeresena Alemsegad anunció el descubrimiento de un niño Au. afarensis de 3.3 millones de años a no 4km de donde fue encontrada Lucy en 1974. No sólo es el fósil juvenil más antiguo jamás encontrado, también es el fósil de homínidos más completo encontrado hasta la fecha. Selam, como el fósil fue apodado (o el bebé de Lucy o el bebé de Dikika), confirmó sugerencias anteriores de que Au. afarensis era bípedo, pero pasaba tiempo en los árboles. La estructura del hombro con sus articulaciones de hombro apuntando hacia arriba y la cresta ósea que corre a lo largo de los omóplatos son como la de los simios, lo que habría facilitado el movimiento arbóreo aunque no fueran tan capaces como los chimpancés de moverse en los árboles.
Los datos paleoecológicos indican que Au. afarensis vivió tanto en pastizales (sabanas) como en entornos boscosos.
Australopithecus africanus
Raymond Dart identificó Australopithecus africanus, “simio sureño africano”, en 1924, el primer australopit reconocido. Este ejemplar era un australopit juvenil, apodado el bebé Taung. Basado en la posición del foramen magnum, Dart afirmó que era un homínido bípedo, pero sus contemporáneos refutaron su afirmación ya que pensaban que el cerebro era demasiado pequeño. La comunidad paleoantropológica no reconoció al bebé Taung como homínido hasta la década de 1950, momento en el que se encontraron otros especímenes fósiles atribuidos a Au. africanus. Au. africanus solo se ha encontrado en el sur de África en sitios como Makapansgat, Sterkfontein y Taung.
.jpg)
Figura\(\PageIndex{6}\) - Australopithecus africanus (Sra. Ples)
Utilizando principalmente métodos biocronológicos ya que Sudáfrica no tiene depósitos geológicos adecuados para otros tipos de técnicas de datación, se ha fechado a 3-2 mya (Scarre 2014). En comparación con Au. afarensis, Au. africanus tiene incisivos más pequeños y molares más grandes; los caninos ya no tienen la apariencia puntiaguda y triangular que se ve en simios y Au. afarensis; sin embargo, Au. africanus todavía exhibe algo de prognatismo en la cara y tiene un cerebro pequeño como el de Au. afarensis. Postcranealmente, Au. africanus conserva algunas características primitivas como las que se ven en Au. afarensis, por ejemplo, brazos más cortos que piernas y forma de mano/pies. Su tamaño también es similar al Au. afarensis. Otra característica derivada son los pilares nasales. Los pilares nasales son un “refuerzo de hueso a cada lado de la abertura nasal de la habilidad en el maxilar superior”, lo que puede ser una adaptación para comer alimentos duros (Becoming Human c.2008). Otros rasgos derivados incluyen un hueso occipital más largo y plano y un hueso frontal más alto, dos rasgos observados en el género Homo. Debido a los rasgos mencionados anteriormente, existe un consenso general de que Au. africanus es descendiente directo de Au. afarensis. Algunos paleoantropólogos sugieren que Au. africanus es el progenitor de Paranthropus robustus (ver abajo). Su relación con el Homo es menos clara.
Australopithecus garhi
Encontrado en Bouri, Etiopía en 1997 por Behane Asfaw y Tim White, Australopithecus garhi es un gracile australopith. “Gracile” se refiere al grosor del hueso; en este caso es menos robusto que sus contemporáneos Paranthropus. Utilizando métodos de datación radioisotópica y biocronométrica, Au. garhi data de 2.5 mya. Se han encontrado pocos especímenes fósiles y los que tienen son relativamente fragmentarios. Se encontraron un cráneo y otros fragmentos de cráneo que sirven como base para la identificación de especies. El tamaño y la forma de sus dientes molares sugieren a algunos investigadores que Au. garhi está relacionado con Paranthropus aethiopicus (ver abajo), pero sus otras características, por ejemplo, el cerebro, la cara y otros dientes, se parecen más al género Homo. Ante esto, algunos investigadores sostienen que es ancestral al Homo.
.jpg)
Figura\(\PageIndex{7}\) - Australopithecus garhi
Australopithecus sediba
Un relativamente recién llegado al escenario, Australopithecus sediba fue encontrado en 2008 en la cueva de Malapa, Sudáfrica por Matthew Berger, el hijo de 9 años del paleoantropólogo principal del proyecto, Lee Berger. Data de 1.95-1.75 mya usando paleomagnetismo, datación de uranio-plomo y biocronología, Au. sediba tiene un mosaico de características que sugieren que puede ser transicional de los australopitos al género Homo, de ahí su nombre “manantial o fuente” en la lengua local de Lesotho. Sin embargo, esta afirmación es polémica ya que las fechas más tempranas para Homo son anteriores a Au. sediba en unos 500,000 años (Becoming Human c2008).

Figura\(\PageIndex{8}\) - Australopithecus sediba
Au. sediba comparte algunos rasgos con Au. africanus, por ejemplo, capacidad craneal, crestas de cejas y cúspides de dientes agrandados (la parte superior puntiaguda de los dientes), lo que hace que algunos afirmen que las dos especies están relacionadas. Sus proporciones de brazo a pierna son igualmente similares a las de los australopitos. Las características que vinculan a Au. sediba con Homo incluyen la forma de la pelvis, el caso cerebral más vertical, los pómulos más pequeños y la forma molar.
Otra controversia surgió recientemente en 2014 cuando Ella Been y Yoel Rak presentaron su estudio de médula espinal (Barras 2014). Afirman que las características de la médula espinal y las mandíbulas de los dos especímenes identificados por Berger son en realidad dos especies diferentes: Homo temprano y un australopith. Adicionalmente, Been y Rak afirman que la colección Malapa contiene los restos de cuatro individuos, no dos. Berger disputa esta afirmación de que hay dos especies presentes. Puedes leer más detalles sobre este debate enNew Scientist [opcional].
Australopitos Robustos
Tres especies robustas de homínidos surgieron en el período Plio-Pleistoceno: Paranthropus aethiopicus, Paranthropus boisei y Paranthropus robustus. Tienen características morfológicas que sugieren que estaban bien adaptados para comer alimentos duros que necesitaban molienda, lo que llevó a que fueran identificados como “robustos”. Sus incisivos y caninos son pequeños, mientras que sus premolares y molares son bastante grandes. El arco cigomático está acampanado, lo que permite músculos masticatorios más grandes que se necesitan para moler alimentos duros. Esto tiende a hacer que la cara sea como un plato-como. Una prominente cresta sagital encabeza el cráneo, otra adaptación masticatoria, ya que el duro músculo temporal es un hueso robusto sobre el que unirse. Se ha argumentado que los robustos australopitos estaban tan bien adaptados que no pudieron responder con la suficiente rapidez a las cambiantes condiciones ambientales, lo que provocó su extinción en 1.0 mya. Recientemente, la evidencia dental de P. boisei sugiere que los alimentos duros no eran la dieta primaria de al menos P. boisei, sino que eran alimentos retrocedidos, o alimentos que se comían cuando los alimentos preferidos no estaban disponibles (Becoming Human c2008).
Nota
Existe debate sobre si las diferencias mencionadas anteriormente califican a los australopitos robustos para estar en un género separado de los australopitecinos. En algunas obras antropológicas se utiliza el género Australopithecus. En otras, como esta obra, se utiliza Paranthropus.
Paranthropus aethiopicus
Paranthropus aethiopicus tiene fecha 2.7-2.3 mya. Si bien fue identificada por primera vez como especie en 1967 por investigadores franceses que encontraron una mandíbula desdentada en Omo, Etiopía, no fue aceptada hasta 1985 cuando Alan Walker y Richard Leakey encontraron el “cráneo negro” en el lago Turkana, Kenia. Se llama cráneo negro porque durante el proceso de fosilización se tiñó de negro por los minerales presentes en el sedimento. Otros fósiles de P. aethiopicus se han encontrado en Laetoli, Kenia desde el descubrimiento de 1985; sin embargo, nunca se ha recuperado material postcraneal.
.jpg)
Figura\(\PageIndex{9}\) - Paranthropus aethiopicus
P. aethiopicus exhibe una mandíbula poderosa, cresta sagital bien desarrollada y molares megadontes o extremadamente grandes. Su capacidad craneal, las articulaciones entre la mandíbula y el cráneo, y la cara prognática son similares a Au. afarensis. La prominencia de la cresta sagital hacia la parte posterior del cráneo también es similar a Au. afarensis. Estas características no están presentes en otros australopitos robustos o Au. africanus. Estas similitudes llevan a algunos paleoantropólogos a afirmar que Au. afarensis es ancestral de P. aethiopicus.
P. aethiopicus comparte varias características con otras especies robustas, como arcos cigomáticos posicionados hacia adelante, una cara ahuecada (ver reconstrucción de P. boisei abajo), huesos gruesos del paladar y grandes premolares y molares.
Paranthropus boisei
Paranthropus boisei comparte varias características con P. aethiopicus, incluyendo una cara abombada, fuerte cresta sagital y grandes premolares y molares (de hecho, los molares son aproximadamente cuatro veces más grandes que los humanos modernos (Smithsonian 2015)). A diferencia de P. aethiopicus, la cresta sagital se posiciona hacia la parte frontal del cráneo en lugar de hacia la parte posterior. Otras características únicas incluyen los premolares con forma de molares, el arco cigomático forma un arco circular, el foramen magnum es corto y en forma de corazón, y el cerebro es grande que los otros australopitos robustos. Poco se sabía sobre las estructuras postcraneales de P. boisei hasta el descubrimiento 2010/11 de fragmentos de restos postcraneales en Olduvai Gorge, Tanzania. Fragmentos del radio, cúbito, fémur y tibia apoyan sugerencias anteriores de que P. boisei era sexualmente dimórfico. Estos restos postcraneales indican que P. boisei estaba “... más robustamente construido, combinando la locomoción bípeda terrestre y algunos comportamientos arbóreos... [tiene] músculos de antebrazos más bien formados que se utilizaron para escalar, manipulación fina y todo tipo de comportamientos” (Sci-News.com 2013).
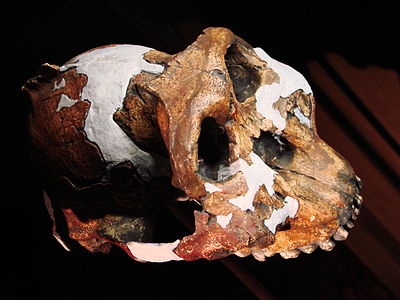.jpg)
Figura\(\PageIndex{10}\) - Paranthropus boisei
P. boisei fue descubierto en 1955 en Olduvai Gorge por Mary Leakey. No fue hasta 1959 que Leakey identificó al fósil como una nueva especie y Zinjanthropus boisei, apodado “Zinj”. Los descubrimientos posteriores de fósiles similares y otros australopitos robustos dieron como resultado un consenso entre los investigadores de que Zinj pertenecía al género Paranthropus. Se han encontrado fósiles de P. boisei en varios sitios de África Oriental, incluyendo Peninj, Tanzania, Konso, Etiopía y Koobi Fora y Turkana Occidental, ambos en Kenia. Vivió en pastizales, pero también áreas boscosas cercanas a fuentes de agua, entre 2.3-1.2 mya, convirtiéndola en una especie de larga vida. No hay evidencia que sugiera que P. boisei sea ancestral de cualquier homínido posterior.

Figura\(\PageIndex{11}\) - Paranthropus boisei; modelo de macho adulto (Museo Smithsonian de Historia Natural)
Paranthropus robustus
Paranthropus robustus es el único australopit robusto que se encuentra en el sur de África. Descubiertos por primera vez en 1938 por Robert Broom en Kromdraii, Sudáfrica, los fósiles de P. robustus también fueron recuperados de Swartkrans y Drimolen, Sudáfrica. Al igual que las otras especies robustas, tiene dientes de mejilla megadonte (premolares y molares), grandes arcos cigomáticos, cara en forma de plato, cresta sagital y un alto grado de dimorfismo sexual. Sus características únicas incluyen un tamaño cerebral mayor que otros australopitos y morfología de la mano, lo que sugiere un agarre capaz de usar herramientas. P. robustus tiene fecha de 2.0-1.2 mya.
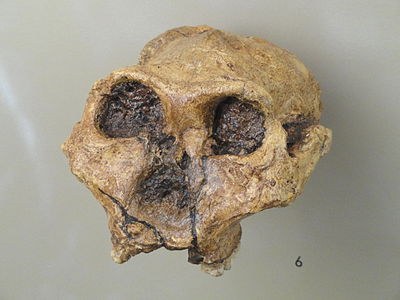
Figura\(\PageIndex{12}\) - Paranthropus robustus
Referencias
- Alemseged Z, Spoor F, Kimbel WH, Bobe R, Geraads D, Reed D, Wynn JG. 2006. Esqueleto juvenil de homínidos tempranos de Dikika, Ethiopoa. Naturaleza 443 (21 Sep): 296-301. Disponible en: http://www.nature.com.offcampus.lib....ture05047.html. doi:10.1038/naturaleza05047.
- Bailey SE. 2006. Sahelanthropus tchadensis. En: Enciclopedia de antropología, Vol. 5. Thousand Oaks (CA): SAGE Referencia. p. 2044-2045.
- Barras C. 2014. Los fósiles humanos del “vínculo faltante” pueden ser un revoltijo de especies. Nuevo Sci [Internet] [citado 2015 Ago 13]; 222 (2964). Disponible en: https://www.newscientist.com/article/mg22229643-200-human-missing-link-fossils-may-be-jumble-of-species/
- Volviéndose Humano. c2008. El linaje humano a través del tiempo. Instituto de Orígenes Humanos [Internet] [citado 2015 Ago 3]. Disponible en: http://www.becominghuman.org/node/human-lineage-through-time
- Domínguez-Rodrigo M, Pickering TR, Baquedano E, Mabulla A, Mark DF, Musiba C, et al. 2013. Primer esqueleto parcial de un Paranthropus boisei de 1.34 millones de años de edad procedente del lecho II, Garganta de Olduvai, Tanzania. PLoS ONE 8 (12): e80347. Disponible en: http://journals.plos.org/plosone/art...l.pone.0080347. doi:10.1371/journal.pone.0080347
- eFósiles [Internet] [citado 2015 ago 10]. Departamento de Antropología, La Universidad de Texas en Austin. Disponible en: http://efossils.org/
- Harmand S, Lewis JE, Feibel CS, Lepre CJ, Prat S, Lenoble A, Boës X, Quinn RL, Brenet M, Arroyo A, Taylor N, Clément S, Daver G, Brugal JP, Leakey L, Mortlock RA, Wright JD, Lokorodi S, Kirwa C, Kent DV, Roche H. 2015. Herramientas de 3.3 millones de años de Lomekwi 3, Turkana Occidental, Kenia. Naturaleza 521 (21 de mayo): 310-315. Disponible en: http://www.nature.com.offcampus.lib....ture14464.html. doi:10.1038/naturaleza14464.
- Caza KD. Australopitecinos. En: Enciclopedia de antropología, Vol. 1. Thousand Oaks (CA): Referencia SAGE, 2006. p.311-317.
- Jurmain R, Kilgore L. Trevathan W. 2013 Esenciales en antropología física. Belmont (CA): Wadsworth Cengage Learning.
- Larsen, CS. 2014. Nuestros orígenes: descubrir la antropología física. Nueva York (NY): W. W. Norton & Company, Inc.
- Lebatard AE, Bouriès DL, Duriner P, Jolivet M, Braucjer R, Carcaillet J, Schuster M, Arnaud N, Monié P, Lihoreau F, Likius A, Macaye HT, Vignaud P, Brunet M. 2008. Datación de nucleidos cosmogénicos de Sahelanthropus tchadensis y Australopithecus bahrelghazali: Homínidos Mio-Pliecenos de Chad. Proc Natl Acad Sci U S A [Internet] [citado 2015 ago 3]; 105 (9): 3226-3231. Disponible en: http://www.pnas.org/content/105/9/3226.full. doi: 10.1073/pnas.0708015105
- Richmond BG, Jungers WL. 2008. Morfología femoral de Orrorin tugenensis y evolución del bipedalismo homínico. Ciencia [Internet] [citado 2015 ago 3]; 319 (5870): 1662-1665. Disponible en: www.jstor.org/stable/20053635
- Scarre C. 2013. El pasado humano. Londres (Reino Unido): Thames & Hudson.
- Sci-news.com [Internet]. 2013 6 dic. Paranthropus boisei: homínido de 1.34 millones de años encontrado en Tanzania. [citado 2015 Ago 12]. Disponible en: http://www.sci-news.com/othersciences/anthropology/science-paranthropus-boisei-hominin-tanzania-01603.html
- Institución Smithsonian [Internet]. 2015 ago 4. ¿Qué significa ser humano? [citado 2015 Ago 11]. Disponible en: http://humanorigins.si.edu/evidence/human-fossils/species
- Su DF. 2013. Los primeros homínidos: Sahelanthropus, Orrorin y Ardipithecus. The Nature Education Knowledge Project [Internet] [citado 2015 Ago 3]. Disponible en: http://www.nature.com/scitable/knowl...hecus-67648286
- Ward CV, Manthi FK, Plavcan JM. 2013. Nuevos fósiles de Australopithecus anamensis de Kanapoi, Turkana Occidental, Kenia (2003-2008). J Hum Evol 65 (5): 501-524. Disponible desde doi:10.1016/j.jhevol.2013.05.006
- Wong K. 2006. Reportaje especial: El bebé de Lucy: sale a la luz un extraordinario nuevo fósil humano. Sci Am [Internet] [citado el 10 de ago de 2015]; 20 de sep. Disponible en: http://www.scientificamerican.com/ar...rt-lucys-baby/