
Abstract
Impact of habitat characteristics and anthropogenic activities on the distribution of macroinvertebrate traits and ecological preferences was assessed in Osun State, Nigerian. Physicochemical parameters involving water depth (WD), channel width (CW), flow rate, discharge, water temperature (W.T), dissolved oxygen (DO), total dissolved solids (TDS), electrical conductivity (EC), pH, biological oxygen demand (BOD), turbidity, nitrate, phosphate and organic matter (OM), were appraised, alongside macroinvertebrates from 5 streams in different geographical locations. Sampling spanned from December 2019 to October 2021, covering early and late dry seasons, as well as early and late rainy seasons. Principal component analysis classified the sites into three impact categories: least impacted (LIS), moderately impacted (MIS), and highly impacted (HIS). Nine traits and ecological preferences displayed significant variations alongside nine physicochemical variables, such as phosphate, OM, WD, discharge, CW, TDS, EC, W.T, and nitrate, within the Fourth-corner figure. Furthermore, these traits and ecological preferences exhibited notable correlations with site impact categories, including–no turbidity preference, spherical body shape, climbers to LIS, preference for transparent water, crawlers, climbers, burrowers, small body size to MIS, and highly tolerant to ocxygen depletion to MIS.This study underscores the effectiveness of macroinvertebrate trait-based approach as a freshwater biomonitoring tool in Osun State, Nigeria, providing essential insights into how environmental factors influence macroinvertebrate traits in lotic systems in the region, with implications not only locally but also for broader eclogical studies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The distribution of macroinvertebrate traits and ecological preferences in tropical streams is strongly influenced by both habitat characteristics and anthropogenic activities (Ngodhe et al. 2014; Edegbene et al. 2019). Physical parameters like water temperature, flow velocity, substrate composition, and depth significantly influence the distribution of macroinvertebrates (Akindele et al. 2022, 2024). Additionally, water quality variables, including DO levels, pH, nutrient concentrations, and pollution, affect their abundance and diversity (Arimoro et al. 2015, 2021). Certain macroinvertebrate species demonstrate varying degrees of tolerance to pollution, making them useful indicators of water quality (Odume et al. 2014). Macroinvertebrate communities respond differently to pollution levels, with pollution-tolerant species dominating degraded sites, while pollution-sensitive species are more abundant in pristine areas (Masese et al. 2009). Moreover, riparian vegetation along stream banks affects macroinvertebrate diversity by offering shade, organic matter, and habitat complexity (Marques et al. 2021; Akinpelu et al. 2024). Thus, human activities contribute to tropical stream ecosystem degradation, impacting macroinvertebrate communities (Jayawardana et al. 2017). Various land use activities, including agriculture, waste mismanagement, grazing, mining, and rising water demand, stress ecosystems (Malmqvist and Rundle 2002; Padgham et al. 2015). Agricultural chemical use degrades water quality through runoff and leaching (Ligeiro et al. 2013; Wantzen and Mol 2013). Mismanagement of domestic and industrial waste also introduce pollutants into water systems, disrupting aquatic health (Qin et al. 2020). Grazing causes riparian deforestation, bank erosion, sedimentation, and nutrient loading, altering benthic animal preferences (Field et al. 2018). Additionally, artisanal mining poses a threat to water quality by discharging waste materials and chemicals, negatively impacting nearby water bodies (Corredor et al. 2021). Largely, human activities near settlements have severe consequences, compromising water quality, diminishing river habitats, and elevating nutrient and pollutant levels (Carvalho et al. 2021; Akindele et al. 2020; 2023). Therefore, these changes result in decreased flow rates, modified hydrological patterns, and transformed compositions of aquatic life, which involve shifts in functional diversity, loss of sensitive species, and the prevalence of more resilient organisms (Freeman et al. 2019).
The taxonomical approach, traditionally employed by ecologists to study the distribution and impact of pollution on freshwater ecosystems, involves identifying and analyzing macroinvertebrates at a taxonomic level, in order to understand their structural patterns (Bo et al. 2017; Ochieng et al. 2019). For well over two decades, there is a growing global interest among scientists in the use of the macroinvertebrate trait-based approach (MTBA) to complement the taxonomic technique in freshwater biomonitoring. MTBA provides broader information on the health of freshwater systems and is less influenced by geographical differences, making it a cost-effective option (Menezes et al. 2010; Desrosiers et al. 2019; Odume 2020; Edegbene et al. 2021). Traits itself, as defined by Odume (2020) and explained by Edegbene et al. (2019), as a quantifiable characteristic that describe an organism’s biology and behavior at an individual level. MTBA is globally popular (e.g., Descloux et al. 2014; Murphy et al. 2017; Desrosiers et al. 2019). Nonetheless in Africa, research using trait-based tools for freshwater biomonitoring is limited (e.g., Odume et al. 2014; Pallottini et al. 2017; Akamagwuna et al. 2019; 2023; Odume 2020). Meanwhile, MTBA, essential for freshwater biomonitoring, is rooted in the Habitat Template Concept (HTC). This underscores the significance of organisms having suitable trait combinations for habitat adaptation, with HTC proposing communication between external structures and traits (Townsend and Hildrew 1994). Unlike traditional taxonomic methods, MTBA offers ecological insights into how macroinvertebrate traits mediate between organisms and their environment (McGill et al. 2006). Although MTBA applications for studying pollution impacts on rivers in Sub-Saharan Africa, particularly Nigeria, are evident (e.g., Edegbene et al. 2019; 2021; Adedapo et al. 2023), there remains a knowledge gap regarding MTBA’s applicability in Nigeria streams.
The ecoregion of Osun State Nigeria presents a favorable environment for utilizing MTBA as an effective tool for evaluating the ecological health of streams and rivers. This region appeal for resettlement is driven by its high demand for water and land, supported by fertile soil and abundant freshwater systems, aligning with the region’s historical agricultural focus (Lasisi et al. 2017). The influx of migrants and indigenous inhabitants, engaging in agricultural and land use activities, has notably impacted regional streams and rivers (Adedeji and Joseph 2018). This has resulted in riparian deforestation, surface runoff of nutrients and pesticides, siltation, changes in aquatic biota structure and function, and modifications in species assemblages (Ashaolu et al. 2019; Aduwo et al. 2023; Akinpelu et al. 2024). The situation is exacerbated by the lack of an effective waste management and treatment plan, leading to uncontrolled discharge of organic pollutants into the water bodies (Onoharigho et al. 2022). The study aimed to assess the reliability of MTBA as a biomonitoring tool in the ecoregion, considering the aforementioned environmental challenges.
Materials and methods
Description of study area
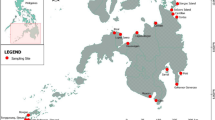
The research encompassed multiple geographic locations in Osun State, Nigeria, specifically Ile-Ife, Ipetumodu, Apomu, Erinjesha, and Apomu. Two sampling stations were chosen in each of the streams (upper reach and lower reach), having a distance of 500m apart (Fig. 1). These streams hold significant ecological importance to both rural and semi-urban communities residing in the surrounding areas, acting as vital assets by providing essential ecosystem services related to water and serving as biodiversity hotspots. The study location falls within the tropical rainforest zone and exhibits two distinct seasons; a dry season from November to March and a wet season from April to October (Omogbai 2010). The average annual rainfall of this location ranges from approximately 1500–2500 mm (59–98 inches), and the average annual temperature is around 27 °C (81 °F) (Ogundari and Ojo 2006). The soil in this region is rich in organic matter, rendering it suitable for agriculture, which serves as the primary and thriving occupation. The soil composition primarily consists of sedentary and metamorphic rocks, which provide ample nutrients such as nitrogen, phosphorus, and potassium (Adeyolanu et al. 2016). However, soil erosion poses a significant challenge in the study area, predominantly caused by riparian deforestation resulting from farming and logging activities, unsustainable agricultural practices like overgrazing, the high rainfall characteristic of the rainforest belt, hilly terrain, and inadequate soil conservation practices. These factors contribute to sedimentation and siltation in numerous streams and rivers within the region (Fasinmirin and Olorunfemi 2014). A Geographical Positioning System (GPS) handset was used to mark out each sampling site in the studied streams, as outlined in Table 1 which contains more detailed information regarding other features of each stream.

Map of Osun State, Nigeria showing locations of the streams and sampling sites. (* indicates upstream station and ** indicates downstream station)
Field sampling, in-situ determinations and environmental variables
Field sampling was conducted from December 2019 to October 2021, encompassing both the early and the peaks of dry and wet seasons. The specific sampling months were as follows: December 2019 (early dry season), March 2020 (late dry season), June 2020 (early rainy season), October 2020 (late rainy season), December 2020 (early dry season), March 2021 (late dry season), June 2021 (early rainy season), and October 2021 (late rainy season). This implies a total of eight sampling periods in a total of 10 stations from five sites. Sampling program was designed to take place within a week per sampling exercise in order to cover all the sites. The vegetation cover and substratum composition were visually assessed (Kennedy and Addison 1987; Addo-Bediako 2021). Water flow rate was determined by measuring the time it took a float to travel a distance of 10 m (Gordon et al. 1994). The depth of the streams was determined using a graduated wooden rod, while the width was measured at the widest point of each station and sampling period using a meter tape (Keke et al. 2020). Various physicochemical water parameters were measured using a digital meter (Multi 3630 IDS, WTW/Xylem Analytics, Germany), including the water temperature (°C), pH, DO (mg L−1), EC (µ Scm−1), and TDS (mg L−1). Additionally, water samples were collected for organic matter (OM) (mg L−1), nitrate (mg L−1) and phosphate (mg L−1) analysis.
Macroinvertebrate sampling
Macroinvertebrate samples were collected concurrently from diverse substrate types, microhabitats (such as vegetation, stones, sand, and gravels), and flow regime zones. A D-frame net with a 500 m mesh size was utilized along a 10 m wade-able stretch in each stream, employing the kick sampling technique. In addition, manual searching was conducted by gathering wood debris and extracting specimens from it. Some macroinvertebrates were observed to be closely associated with rocky substrates and trailing plants, and were collected by carefully examining these microhabitats (Arimoro et al. 2021; Akindele et al. 2022). For each upstream and downstream location, all collected samples representing different biotopes in each station were combined into a single sample representing that of upstream or downstream station. Subsequently, these samples were preserved in 70% ethanol and transported to the laboratory for further analysis. Identification guides by Merritt and Cummins (1996), Day et al. (2003), and De Moor et al. (2003) were utilized as references during the study. Owing to the absence of comprehensive identification guides for macroinvertebrate traits at the species level within Nigerian lotic ecosystems, coupled with constrained taxonomic proficiency, the ascertained traits of macroinvertebrates were confined to the family level (Akamagwuna et al. 2019; Edegbene et al. 2020) (Table 2). It should be noted that, the term “constrained taxonomic proficiency” regarding species trait identification, suggests that, tools developed at the species level may not be widely applicable within Osun State ecoregion of Nigeria. Consequently, previous studies have also employed macroinvertebrate trait-based analysis at the family level of taxonomic resolution (e.g., Ding et al. 2017; Forio et al. 2018; Edegbene et al. 2019).
Selection of macroinvertebrates traits and ecological preferences
A total of 32 attributes, representing seven traits and ecological preferences, were carefully selected for this study (Table 3). To account for the variability and flexibility of these traits and ecological preferences among members of the same macroinvertebrate family, as well as different life-history stages of macroinvertebrates, a fuzzy coding method was employed (Chevenet et al. 1994). These attributes encompassed various aspects such as respiration (tegument, spiracles, and other apparatus), body armouring (hard shell, total sclerotized, partly sclerotized, soft and exposed, and soft and exposed), turbidity preference (silt, opaque, transparent, and no preference), body shape (streamlined, flattened, spherical, and cylindrical), mobility (climber, crawler, sprawler, swimmer, skater, and burrower), body length (ranging from very small (< 5 mm) to very large (> 40–80 mm), and sensitivity to oxygen depletion (highly sensitive, moderately sensitive, moderately tolerant, and highly tolerant). Selection of these traits and ecological preferences was based on their perceived mechanistic relationship with the nature of the habitat as well as prevailing anthropogenic activities observed in the study area. Information on these traits and ecological preferences was primarily extracted from Odume et al. (2018), in conjunction with several other applicable data, (e.g., Kuzmanovic et al. 2017; Desrosiers et al. 2019; and Edegbene et al. 2019). Appendices A.1 and A.2 present the level of affinity for each trait attribute investigated, using Fuzzy codes. In this coding system, “0” signifies no affinity, “1” indicates low affinity, “2” represents moderate affinity, and “3” indicates maximum affinity to the specific trait or ecological preference for the macroinvertebrate family being considered.
Calculation, analyzing and interpreting statistical data
Characterizing the varying levels of impact across the selected rivers
The 10 sites in the 5 stream systems that were studied were divided into three categories based on the levels of impact they experienced: least impacted sites (LIS), moderately impacted sites (MIS), and highly impacted sites (HIS). This categorization was determined through principal component analysis (PCA), which involved analyzing the sites and the physical and chemical factors associated with them (see Table 4). To assign these categories, we first extracted the coordinates of the river sites on the first PCA axis. Then, we calculated the distances between the sites by subtracting the score of the LIS from the score of the HIS. These distances were converted into percentages and a percentile distribution was used to classify the sites into one of the three impact categories. Specifically, sites falling within the 100th to 90th percentile were designated as least impacted, those falling below the 90th to 50th percentile were classified as moderately impacted, and sites below the 50th percentile were considered highly impacted. This methodology was adapted from previous studies by Odume et al. (2016) and Edegbene et al. (2020) who used similar approaches to classify river sites based on physicochemical variables along the first axis of PCA. The specific data for the physicochemical variables used in this study can be found in Appendix B. The PCA analysis was conducted using the vegan package version 2.5.4 in R-statistics (Oksanen et al. 2019).
The RLQ ordination plots depict the interrelationships among physicochemical parameters, macroinvertebrate traits/ecological preferences, and the sampling sites
The spatial distribution of the selected traits and ecological preferences was explored by using a multivariate analysis technique called RLQ, which was developed by Dolédec et al. (1996). RLQ typically combines three datasets: environmental variables (R), taxa (L), and traits (Q). In our case, RLQ was used to integrate physicochemical variables (R), macroinvertebrate taxa traits and ecological preferences (L), and the sampling sites (Q). By utilizing the RLQ ordination plane, we were able to visually analyze how potential traits and ecological preferences were distributed in response to habitat characteristics and human activities. To determine the statistical significance of the RLQ axes, a Monte Carlo test with 999 permutations was conducted at a significance level of p = 0.05. The RLQ analysis was performed using the Ade4 package for R-statistics version 2.5.4 in the R programming platform (Oksanen et al. 2019).
Establishing the connection between specific macroinvertebrate traits and ecological preferences with physicochemical variables
We utilized the fourth-corner analysis, a multivariate technique, to investigate the relationship between physicochemical variables and the selected traits and ecological preferences. This analysis simultaneously evaluates the statistical significance of multiple biological traits, ecological preferences, and physicochemical variables. An advantage of the fourth-corner analysis is its ability to reveal the strength and direction of correlations between traits and physicochemical variables. In this study, macroinvertebrate traits and ecological preferences associated with LIS or MIS on the RLQ ordination plane, and that is/are also exhibited significant positive correlations with DO or significant negative correlations with variables such as EC, TDS, phosphate, nitrate, temperature, BOD, or organic matter, are considered sensitive to anthropogenic activities. Conversely, traits and ecological preferences associated with HIS on the RLQ ordination plane, that equally displayed statistically significant positive correlations with variables like BOD, EC, TDS, nitrates, temperature, organic matter and phosphates, or significant negative correlations with DO, are identified as anthropogenic tolerant traits in a perturbed site (Odume et al. 2016; Edegbene et al. 2020). Ade4 package for R-statistics version 2.5.4 in the R programming platform (Oksanen et al. 2019), was used to perform the fourth-corner analysis.
Results
Impact categories and correlations of data sets on the RLQ
Based on the impact categories presented in Table 4, the following designations was assigned to the respective stations: Olumirin station-1 [OLM-1 (7)] and Olumirin station-2 [OLM-2 (8)] were categorized as LIS. Mogimogi station-1 [MOG-1 (5)], Awoosun station-1 [AWS-1 (3)], Awoosun station-2 [AWS-2 (4)], Abirunmu station-1 [ABR-1 (1)], and Abirunmu station-2 [ABR-2 (2)] were classified as MIS. Furthermore, Aboto station-1 [ABT-1(9)], Aboto station-2 [ABT-2(10)], and Mogimogi station-2 [MOG-2 (6)] were identified as HIS. Figures 2, 3, and 4 indicate a comprehensive depiction of the relationship between physicochemical parameters, macroinvertebrate traits and ecological preferences, as well as different stream sites. This visualization, using RLQ ordination plots, seamlessly combines three sets of data, as each set of data is shown in Fig. 2 (physicochemical and environmental variables-(R)), Fig. 3 (macroinvertebrate taxa traits and ecological preferences (L)), and Fig. 4 (the sampling sites (Q)). Table 5 presents the eigenvalues of Axes 1 and 2, which were 1.454 and 0.397, respectively. The RLQ ordination explained a projected total inertia of 4.65. Axis 1 accounted for 75.75% of the variance of the RLQ for the PCA, while Axis 2 explained 10.76%. The Hill-smith transformed variance for the macroinvertebrate traits and ecological preference were 30.76 and 30.68 for Axis 1 and 2.

R-Ordination plots showing the physicochemical parameters of the sampled streams. R physicochemical parameters: BOD biological oxygen demand, Dch discharge, DO dissolved oxygen, Dep depth, EC electrical conductivity, Flw flowrate, N nitrates, OM organic matter, P phosphate, TDS total dissolved solids, Tep water temperature, Tud turbidity, Wdt width

L-Ordination plots showing the macroinvertebrates traits and ecological preferences. T1 gills, T2 tegument, T3 spiracle, T4 other apparatus, T5 hard shell, T6 total sclerotized, T7 partly sclerotized, T8 soft and exposed, T9 existing in case, T10 silt, T11 opaque, T12 Transparent, T13 no preference, T14 streamlined, T15 flattened, T16 spherical, T17 cylindrical, T18 climber, T19 crawler, T20 sprawler, T21 swimmer, T22 skaters, T23 burrower, T24 very small (< 5 mm), T25 small (> 5–10 mm), T26 medium (> 10–20 mm), T27 large (> 20–40 mm), T28 very large (> 40–80 mm), T29 highly sensitive to oxygen depletion, T30 moderately sensitive to oxygen depletion, T31 moderately tolerant to oxygen depletion, T32 highly tolerant to oxygen depletion

Q-Ordination plots showing the sampled streams in Osun State, Nigeria. ABR-1 (1) abirunmu station-1, ABR-2 (2) abirunmu station-2, AWS-1 (3) awoosun station-1, AWS-2 (4) awoosun station-2, MOG-1 (5) Mogimogi station-1, MOG-2 (6) mogimogi station-2, OLM-1 (7) Olumirin station-1, OLM-2 (8) Olumirin station-2, ABT-1 (9) aboto station-1, ABT-2 (10) aboto station-2
In Figs. 2, 3, and 4, RLQ plots reveal correlations involving certain macroinvertebrate traits and ecological preferences, specifically, (T6) total sclerotized, (T7) partly sclerotized, (T9) existing in case, (T10) turbidity preference for silt, (T13) no preference for turbidity, (T14) streamlined body, (T15) flattened body, (T18) climbing, (T21) swimming, (T26) medium body size (> 10–20 mm), (T27) large body size (> 20–40 mm), (T28) very large body size (> 40–80 mm), (T29) highly sensitive to oxygen depletion, and (T30) moderately sensitive to oxygen depletion with some physicochemical variables involving water discharge, channel width, DO, flow rate, pH, and BOD, in “LIS”. On Axis 2 of the RLQ in Figs. 2, 3, and 4, gills (T1), tegument (T2), spiracle (T3), transparent body (T12), cylindrical body shape (T17), crawling (T19), sprawling (T20), skating (T22), burrowers (T23), small body size (T25), and moderate tolerance to oxygen depletion (T31) were all associated with phosphate concentration in “MIS” while other apparatus (T4), hard shell (T5), soft and exposed body (T8), preference for opaque water (T11), very small body length (< 5 mm) (T24), and high tolerance to oxygen depletion (T32) are associated with water depth, turbidity, nitrates, temperature, EC and TDS in"HIS” (Figs. 2, 3, and 4).
The significant macroinvertebrate traits/ecological preferences and the physicochemical parameters in the sampled streams using Fourth-corner test
Figure 5 displays the Fourth-corner test results, revealing significant correlations between physicochemical characteristics and the traits/ecological preferences of sampled stream macroinvertebrates. In the context of the analysis, red cells signify significant positive relationships, blue cells indicate significant negative correlations, and gray cells denote non-significant relationships in the table. According to the Fourth-corner test results, there was a significant positive correlation exists between Tegument respiration (T2) and phosphate concentration. Also, a significant negative correlation was observed between Transparency (T12) and OM. In contrast, “No turbidity preference (T13)” was significantly inversely correlated with water depth. Additionally, “Spherical (T16)” and “climbers (T18)” exhibited a positive correlation with water discharge. Moreover, “crawlers (T19) recorded negative correlation with channel width. Thus, macroinvertebrate “burrowers (T23)” significantly positively correlated with three pollution indicating variables involving TDS, EC, and phosphate. Lastly, small-bodied macroinvertebrates (> 5–10 mm) exhibited a significant positive correlation with phosphate concentration, and finally, macroinvertebrates with a high tolerant to oxygen depletion exhibited a significant positive correlations with both the temperature and nitrate concentration. The highest abundance of certain insect families in the LIS was observed among the Ephemeroptera (Baetidae, Heptageniidae, Leptophlebiidae, and Oligoneuridae), Plecoptera (Perlidae), and Trichoptera (Hydrophilidae) groups. These families possess specific characteristics and ecological preferences, such as, gill respiration, soft and exposed body armor, a preference for silt, streamlined, flattened, and cylindrical body shapes. They exhibit mobility through climbing, crawling, and swimming, and have small to medium-sized bodies. Furthermore, these insects show high to moderate sensitivity to low oxygen levels. On the other hand, the HIS recorded the lowest abundance for these insect families (Table 2).

Fourth-corner test performed for macroinvertebrates traits/ecological preferences and analyzed physicochemical variables. Red indicates significant positive correlation (P ≤ 0.05) and blue indicates significant negative correlation (P ≤ 0.05). Traits T2 tegument, T12 transparent, T13 no preference, T16 spherical, T18 climber, T19 crawler, T23 burrower, T25 small body size (> 5–10 mm), T32 highly tolerant to oxygen depletion (color figur online)
Discussion
The study underscores that macroinvertebrate traits in stream habitats are shaped by habitat characteristics and human impacts, such as deforestation for farming, and grazing activities, poor domestic waste management, abattoir services, and water abstraction. These observations align with Diaz et al. (2007), Menezes et al. (2010), Li et al. (2012), and recent works by Odume (2020), Edegbene et al. (2020, 2021) and Akinpelu et al. (2024). RLQ ordination analysis distinguishes anthropogenic-sensitive traits and ecological preferences of macroinvertebrates at LIS from other impact categories stream sites (i.e., MIS and HIS). The association of body armor, turbidity preference, body shape, modes of mobility, body length and sensitivity to oxygen depletion traits with LIS aligns with the findings of Edegbene et al. (2021), as several of these traits and ecological preferences were linked with near-natural sites and slightly disturbed sites, which correspond to LIS and MIS in this study. The sensitive traits and ecological preference that were significantly influenced by habitat features and anthropogenic activities were- preference for no turbidity and climbing, as both were associated with LIS on the RLQ ordination. Trait preference for no turbidity was significantly inversely correlated with water depth, while climbing trait was positively correlated with water discharge. This contrasts with Edegbene et al. (2020), who reported large body size as a LIS sensitive trait positively correlated with DO. Divergent outcomes regarding the sensitive trait may stem from differences in abiotic conditions among freshwater habitats and the trait syndrome under examination. The association of climbing trait with LIS may be due to the ability of the animals to navigate and colonize habitats with complex structures, such as rocks, logs, and vegetation, which characterize the LIS. However, the significant correlation of climbing trait with water discharge on the Fourth-corner is expected. According to Merritt and Cummins (1996), macroinvertebrate with climbing ability are often found in habitats having clean water flow and complex physical structures needed for climbing such as vegetation, rocks and wood debris, offering essential surfaces for attachment and movement, enabling the animals to feed, reproduces and effectively avoid predators. In this study, LIS represents Olumirin stream—an undisturbed environment, with complex biotopes and free from anthropogenic activities. In the RLQ analysis, a notable connection emerged between larger-bodied macroinvertebrates and the LIS, aligning with the habitat template concept (HTC) proposed by Townsend and Hildrew (1994) and Poff et al. (2006), which posits that organisms with larger body sizes often inhabit less impacted sites. However, contrary evidence from Statzner and Beche (2010), Odume et al. (2014), and Odume (2020) associates large body size macroinvertebrates with impacted sites. The observed association of this trait with LIS on RLQ in this study can be ascribed to ability to reduce surface area-to-volume ratio, facilitating rapid diffusion of toxicants and rendering them less susceptible to contamination. Edegbene et al. (2021) similarly explain that the morphological structure (large body size) provides a protective buffer against contamination. Differing from Odume (2020), who linked macroinvertebrate swimmers to highly polluted sites, this study uncovers a unique association of this trait with LIS. Variation in the results can be linked to streamlined body form of the animals, high concentration of DO, and their predatory strategies. Vogel (2008), mentioned that, macroinvertebrate swimmers (e.g., beetles and bugs) possess streamlined bodies and specialized appendages enhancing swimming efficiency by reducing drag in turbulent water. Xu et al. (2014) also noted that the ability to swim in well-oxygenated environments provides necessary energy for navigating obstacles in the environment and sudden human perturbations. Martínez and Castro (2007) added that swimming trait offers advantages to active predator beetles and bugs, like the Hydrophilidae family, aiding in effective prey search and capture. Moreover, beetles and bugs have evolved structures enabling them to carry air bubbles or store atmospheric oxygen for respiration when submerged, especially during periods of high water (Jones 2019). Remarkably, the LIS in this study is a highly intricate habitat, characterized by the highest DO concentration and year-round water discharge. This explains the observed association of swimming trait in some macroinvertebrate families (e.g., Dytiscidae, Hydrophilidae, Psephenidae, Baetidae, Leptophlebiidae, and Nepidae) with the site. Gill-utilizing macroinvertebrates are increasingly susceptible to clogging in polluted freshwater (Akamagwuna et al. (2019). In contrast, our study not only highlights ecological distinctions between MIS and HIS but also demonstrates MIS lower vulnerability to this trait. The association of gill-trait to MIS can be linked to several factors involving niche partitioning, hydrodynamic condition of the site group, as well as physiological plasticity (Reynolds et al. 2013). The association of tegument, cylindrical body shape and moderately sensitive to oxygen depletion traits to MIS is anticipated. According to Desrosiers et al. (2019), cutaneous organisms are less sensitive to oxygen depletion, thriving in disturbed sites. Cylindrical macroinvertebrates, with their generalist feeding habits and burrowing capabilities, utilize their body shape to exploit organic matter in polluted environments (Akamagwuna et al. 2022). Locomotive traits such as crawling, sprawling, skating, and burrowing are associated with MIS and their distribution aligns with elevated phosphate concentration. This contrasts Edegbene et al. (2021) finding linking these traits to near-natural stations which corresponds to LIS in this study. This scenario potentially indicate the adaptive capabilities of these animals to moderately impacted flowing systems (Pandey et al. 2022). Small body size is resilient to pollution, aligning with Townsend and Hildrew’s (1994) prediction and assertions of Castro et al., (2018) and Edegbene et al., (2023) that rapid reproduction confers resistance to pollution. Macroinvertebrates utilizing alternative respiratory structures, like elytra, spiracles, and lungs, exhibit clear associations with HIS, aligning with Statzner and Bêche (2010) perspective on specialized respiration aiding oxygen acquisition in a low DO environment. In this study, this adaptive trait reduces dependence on DO concentration, potentially influenced by warmer temperatures in riparian deforested HIS. Diaz et al. (2007) noted the prevalence of aerial-respiring Coleoptera and Diptera in polluted settings. Also in our investigation, the prevalence of Coleoptera (e.g., Dytiscidae), Diptera (e.g., Chironomidae), Sorbeoconcha (e.g., Thiaridae), and Venerida (e.g., Cyrenidae) in HIS could be linked to augmented respiration through additional structures. The anticipated correlation of highly tolerant to oxygen depletion trait, preference for opaque water, and soft and exposed body with HIS aligns with increased nutrients, temperature, EC, and TDS. Similar findings were reported by Akamagwuna et al. (2019), Edegbene et al. (2021) and Edegbene et al. (2023).
This study equally demonstrated noteworthy links between nine physicochemical parameters and nine biological traits and ecological preferences. Some trait categories showed positive or negative correlations with anthropogenic-indicating physicochemical parameters, as depicted in the Fourth-corner figure. The observed positive correlation between tegumental macroinvertebrates and phosphate concentration in MIS can be attributed to provision of food resource and availability of DO. Elevated phosphate levels in water systems, often from agricultural fertilizer runoff, boost algae growth, a key food source for macroinvertebrates (Mwedzi et al. 2016). Algal blooms increase photosynthetic activity, elevating oxygen levels in water, benefiting tegument-dependent macroinvertebrates, as observed in MIS. This aligns with Statzner and Bêche’s (2010) result, emphasizing tegumental adaptation in macroinvertebrates for nutrient extraction and protection from high phosphate levels. In contrast to Edegbene et al. (2021) who reported tegument respiration in heavily impacted sites, this study observed association of Planorbidae and Gyrinidae with MIS, an algae-rich environment fostered by phosphate concentration. Nevertheless, the result is consistent with Edegbene et al. (2020) and Mohammed et al. (2021), who reported positive correlations between Gyrinidae/Planorbidae and phosphate concentration. Also, previous studies (e.g., Lamouroux et al. 2004; Tomanova and Usseglio-Polatera 2007; Tomanova et al. 2008; Desrosiers et al. 2019) confirm tegument-respiring macroinvertebrates’ resilience to low DO levels, thriving in disturbed environment. The correlation between macroinvertebrates preferring “transparent” water and organic matter (OM) in MIS reveals an inverse relationship, that is; higher OM corresponds to lower abundance of these macroinvertebrate traits. This aligns with expectations, as elevated OM from agricultural runoff and human activities can diminish water clarity, making these animals more susceptible to predation, limiting their food access, and reducing DO levels in the aquatic ecosystem (Edegbene et al. 2019). Consequently, increased OM in lotic systems may intensify competition for limited food resources, favoring more tolerant species (Friberg et al. 2010). The negative correlation between macroinvertebrate with “No turbidity preference” and water depth in LIS suggests that, these animals are less abundant at greater depths. This trend could be attributed to the physical constraints these animals face when adapting to different water depths. Many macroinvertebrates are specifically adapted to thrive in shallow waters (Leslie and Lamp 2017), utilizing various means of mobility such as crawling, climbing, and burrowing within their habitats. Conversely, they could be more susceptible to predators in deeper waters where they have limited access to food with accompanied decrease in oxygen level and light penetration, which further restricts their survival capabilities. Macroinvertebrate crawlers showed inverse correlation with channel width, as indicated on the Fourth-corner figure. Thus, on the RLQ, crawlers were associated with MIS while channel width associated with LIS. Basically, this observation implies that, the wider the channel width; the lower the abundance of crawlers. As observed, the wider channel of LIS (Olumirin site) is suggesting to be less favorable to macroinvertebrate crawlers, in their search for food, shelter, and protection. Though, streams with wider channels provide diverse habitats for macroinvertebrates to thrive (Arimoro et al. 2011), non-crawling species could outcompete their crawling counterparts, leaving them more vulnerable to various risks. This result opposes a previous report by Edegbene et al. (2021), that reported macroinvertebrate crawlers in near-natural sites. This inconsistency in observations could be attributed to variations in sites topography and habitat characteristics of the study locations. Macroinvertebrate “climbers” and those with a “spherical” body shape in LIS exhibit a positive correlation with water discharge. This connection with climbers is attributed to specialized structures like hooks or claws, allowing them to cling to stable substrates such as submerged rocks and wood debris in LIS. This enables climbers to actively forage for food even during high-water flow (Yetter 2012). This correlation parallels Edegbene et al. (2021) findings, associating climbers with minimally impacted sites. The positive link between “spherical” body shape and water discharge in LIS could be credited to unique adaptations, as seen in Ampullariidae. This adaptation could be responsible for the stabilization of this animal in fast-flowing waters, aiding efficient oxygen uptake and access to food resources.
The anthropogenic-sensitive trait in this study is “preference for transparent water”. This trait is associated with MIS in the RLQ and shows a significant inverse relationship with organic matter in the Fourth-corner figure. Additionally, anthropogenic tolerant trait and ecological preference is “highly tolerant to oxygen depletion”. Besides being associated with HIS in the RLQ, it was significantly positively correlated with temperature and nitrates in the Fourth-corner. This observation contradicts Edegbene et al. (2020), who reported preference for coarse particulate organic matter (CPOM) and “very small body size” as the sensitive signature traits in heavily impacted site. The inverse relationship between macroinvertebrate families with preference for transparent water, such as Calopterygidae, Chlorocyphidae and Perlidae and organic matter in MIS could be linked to the sensitivity of the animals to elevated organic matter as these animals prefer clearer water for feeding, respirartion and reprocuction (Cummins, 1974). Likewise, the positive correlation observed between highly tolerant to oxygen depletion macroinvertebrates (e.g., Chironomidae, Nepidae, Belostomatidae, Ampullariidae, Gyrinidae, and Iridinidae) and water temperature in HIS, can be attributed to their unique adaptations, including resistance to thermal stress due to their hard shell, efficient respiratory systems facilitated by their soft and exposed bodies, and a very small body to volume ratio. Interestingly, macroinvertebrate families found in MIS and HIS have these traits. Odume et al. (2012) and Dabessa et al. (2021) found that certain macroinvertebrate families, like Chironomidae, possess a remarkable ability to transport oxygen, an adaptation that enables them to survive in environments with insufficient oxygen levels. This study underscores mobility’s significance, highlighting a notable connection between macroinvertebrate burrowers and pollution indicators, such as EC, TDS, and phosphate. The association suggests burrowers contribute to ion and dissolved solids release, serving as vital nutrients. Williams-Subiza and Brand (2021) noted that high TDS and EC levels in streams support more organic matter, a primary food source for burrowers. Consequently, in HIS, impacted by anthropogenic activities, substantial organic inputs are evident, explaining the positive correlation of burrowers with the site. The positive correlation between burrowers and phosphate in MIS may be connected to maintainers of burrows and finding stable habitats common to species like Planariidae and Lumbricidae. Bendary et al. (2023) reported the domination of macroinvertebrate burrowers in elevated phosphate environment. “Very small body length”, as well as a “soft and expose body” macroinvertebrates are correlated with HIS on the RLQ. This aligns with Edegbene et al. (2020). Furthermore, the positive significant correlation between “small body length” macroinvertebrates and phosphate concentration in MIS on the Fourth-corner can be attributed to capacity for rapid reproduction and efficient nutrient extraction, facilitated by their higher surface area to volume ratio common to some macroinvertebrate families including Veliidae, Notonectidae, Naucoridae, Gerridae, Baetidae, Leptophlebiidae, and Gyrinidae (Edegbene et al. 2021). On the contrary, Odume (2020) linked “small body length” to highly polluted site in this study, the association of this trait to MIS could be connected to ability of these animals to extract essential nutrients from their surrounding through filtration. The positive correlation observed between tolerant to oxygen depletion macroinvertebrates and nitrate in HIS can be attributed to nutrient availability and eutrophication resulting from organic matter loads. Kilonzo et al. (2014) and Wurtsbaugh et al. (2019) opined that elevated nitrate levels in lotic systems often arise from the application of fertilizers on farmlands within the watersheds. Based on the observations in this study, macroinvertebrates capable of tolerating low DO conditions caused by anthropogenic activities, may benefit from the increased availability of food and reduced competition among other species. This helps to explain the co-occurrence of macroinvertebrate families, including Iridinidae, Cyrenidae, Ampullariidae, Thiaridae, Belostomatidae, Nepidae, Chironomidae, and Lumbricidae, in HIS in this study. The highest abundance of Ephemeroptera, Plecoptera and Trichoptera (EPT) families with associated traits and ecological preferences in LIS can be attributed to factors like good water quality, suitable substrate (e.g., rocks, gravel, and sand), and nutrient availability from riparian vegetation. Conversely, the lowest family abundance observed at HIS is credited to severe anthropogenic activities in the site group (Akindele et al. 2020, 2022; Arimoro et al. 2021). This observation also underscores the role of habitat characteristics and anthropogenic activities in influencing the distribution of macroinvertebrate traits in lotic ecosystems.
Conclusion
This study underscores the ecological significance of macroinvertebrates traits in responding to environmental conditions and human-induced disturbances in aquatic ecosystems. The occurrence and distribution of these traits in lotic systems provide crucial insights, serving as indicators of ecological health. Nevertheless, the study emphasizes the importance of considering physical factors (vegetation, water depth, channel width, water discharge) and ecological indicators (DO, temperature, TDS, EC, nitrate, phosphate, OM) when exploring the connection between macroinvertebrate traits and the environment. This research identifies two specific sensitive and tolerant traits and ecological preferences—preference for clear/transparent water, and highly tolerance to oxygen depletion—emphasizing their correlation with habitat features and anthropogenic activities. These findings suggest the potential applicability of MTBA in other regions of Nigeria, advocating for increased investment in taxonomy and further exploration of aquatic biomonitoring using MTBA. Ultimately, this study lays the groundwork for MTBA, marking the first of its kind in Osun State, Nigeria.
Data availability
All the data relevant to this study are readily available on request at Department of Zoology, Hydrobiology and Conservation Unit, Obafemi Awolowo University, Ile-Ife, Osun State.
Code availability
Not applicable.
References
Addo-Bediako A (2021) Spatiotemporal distribution patterns of benthic macroinvertebrate functional feeding groups in the Blyde River, South Africa. Appl Ecol Environ Res 19(3):2241–2257
Adedapo AM, Kowobari ED, Fagbohun IR, Oladeji TA, Amoo TO, Akindele EO (2023) Using macroinvertebrate functional traits to reveal ecological conditions of two streams in Southwest Nigeria—a case study. Aquat Ecol 57:1–17
Adedeji JA, Joseph AF (2018) Urbanisation forces on the landscapes and the changing value-systems of Osun Sacred Grove UNESCO Site, Osogbo, Nigeria. Landsc Res 43(6):798–816
Adeyolanu OD, Ogunkunle AO, Moral MT (2016) Comparison of qualitative and quantitative approaches to soil quality assessment for agricultural purposes in South-western Nigeria. Cogent Food Agric. https://doi.org/10.1080/23311932.2016.1149914
Aduwo I et al (2023) Assessment the impact of Anthropogenic activities on sediment physico-chemical characteristics of Owalla Reservoir, Osun State, Southwest, Nigeria. https://doi.org/10.21203/rs.3.rs-2475475/v1
Akamagwuna FC, Mensah PK, Nnadozie CF, Odume ON (2019) Evaluating the responses of taxa in the orders Ephemeroptera, Plecoptera and Trichoptera (EPT) to diment stress in the Tsitsa River and its tributaries, Eastern Cape. S Afr Environ Monit Assess 191:664
Akamagwuna FC, Odume ON, Richoux NB (2022) Exploring the community structure of Afrotropical macroinvertebrate traits and ecological preferences along an agricultural pollution gradient in the Kat River, Eastern Cape. S Afr Ecol Indic 135:108570
Akamagwuna FC, Odume ON, Richoux NB (2023) Agricultural disturbance affects taxonomic and functional diversity of Afrotropical macroinvertebrate composition in a South African river system. Environ Sustain Indic 18:100251
Akindele EO, Omisakin OD, Oni OA, Aliu OO, Omoniyi GE, Akinpelu OT (2020) Heavy metal toxicity in the water column and benthic sediments of a degraded tropical stream. Ecotoxicol Environ Saf 190:110153. https://doi.org/10.1016/j.ecoenv.2019.110153
Akindele EO, Adedapo AM, Fagbohun IR, Akinpelu OT, Amoo TO, Aliu OO, Adeniyi AV (2022) Conservation evaluation of three Nigerian streams in different zones demonstrate why pristine freshwater ecosystems in the Afrotropics should be protected. Aquat Conserv 32(4):702–709
Akindele EO, Adedapo AM, Fagbohun IR, Akinpelu OT, Aliu OO, Kowobari ED (2023) Macroinvertebrate metric indicators should be juxtaposed with the community conservation index as ecological tools for conservation evaluation of pristine freshwater ecosystems. Biologia. https://doi.org/10.1007/s11756-023-01315-8
Akindele EO, Adedapo AM, Akinpelu OT, Fagbohun IR, Kowobari ED, Oladeji TA et al. (2024) Freshwater macroinvertebrates along the Nigeria-Cameroon border enhance the conservation value of the lower Guinea forest biodiversity hotspot. J Environ Manag 355:120532. https://doi.org/10.1016/j.jenvman.2024.120532
Akinpelu OT, Arimoro FO, Ayanwale AV, Chukwuemeka VI, Adedapo AM, Akindele EO (2024) Comparative investigation provides further insights on how riparian deforestation and different land use impact the distribution of macroinvertebrates in Nigeria streams. Ecohydrology Hydrobiol. https://doi.org/10.1016/j.ecohyd.2024.01.006
Arimoro FO, Ikomi RB, Ajuzieogu IO, Nwadukwe FO (2011) Temporal and spatial variability in macroinvertebrate community structure in relation to environmental variables in Ajijiguan Creek, Niger Delta, Nigeria. Afr J Aquat Sci 36(1):57–66
Arimoro FO, Odume NO, Uhunoma SI, Edegbene AO (2015) Anthropogenic impact on water chemistry and benthic macroinvertebrate associated changes in a Southern Nigeria stream. Environ Monit Assess 187:1–14
Arimoro FO, Abubakar MD, Obi-Iyeke GE, Keke UN (2021) Achieving sustainable river water quality for rural dwellers by prioritizing the conservation of macroinvertebrates biodiversity in two Afrotropical streams. Environ Sustain Indic 10:100103
Ashaolu ED, Jacob FO, Ifatokun PI (2019) Assessing the spatio-temporal pattern of land use and land cover changes in Osun drainage basin, Nigeria. J Environ Geogr 12(1–2):41–50
Bendary RE, Goher ME, El-Shamy AS (2023) Taxonomic and functional diversity of macroinvertebrates in sediment and macrophyte habitats: a case study, the Ibrahimia Canal, Nile River, Egypt. Egypt J Aquat Res 49(2):129–135
Bo T, Alberto D, Francesca B, Stefano F, Alex L (2017) Biomonitoring with macroinvertebrate communities in Italy: what happened to our past and what is the future? J Limnol 76(s1):21–28. https://doi.org/10.4081/jlimnol.2016.1584
Carvalho FG, Duarte L, Nakamura G, Seger GDS, Juen L (2021) Changes of phylogenetic and taxonomic diversity of Odonata (Insecta) in response to land use in Amazonia. Forests 12(8):1061
Castro DMP, Dolédec S, Callisto M (2018) Land cover disturbance homogenizes aquatic insect functional structure in neotropical savanna streams. Ecol Indic 84:573–582. https://doi.org/10.1016/j.ecolind.2017.09.030
Chevenet F, Dolédec S, Chessel D (1994) A fuzzy coding approach for analysis of long-term ecological data. Freshw Biol 31:295–309
Corredor JAG, Pérez EH, Figueroa R, Casas AF (2021) Water quality of streams associated with artisanal gold mining; Suárez, Department of Cauca. Colombia Heliyon 7(6):e07047
Dabessa M, Lakew A, Devi P, Teressa H (2021) Effect of environmental stressors on the distribution and abundance of macroinvertebrates in upper awash river at Chilimo Forest, West Shewa, Ethiopia. Int J Zool. https://doi.org/10.1155/2021/6634168
Day JA, Harrison AD, De Moor IJ (2003) Guides to the freshwater invertebrates of Southern Africa: Diptera. Water Research Commission Pretoria
Descloux S, Datry T, Usseglio-Polatera P (2014) Trait-based structure of invertebrates along a gradient of Sediment colmation: Benthos versus hyporheos responses. Sci Total Environ 466–467:265–276
Desrosiers M, Usseglio-Polatera P, Archaimbault V, Larras F, Methot G, Pinel-Alloul B (2019) Assessing anthropogenic pressure in the St. Lawrence River using traits of Benthic macroinvertebrates. Sci Total Environ 649:233–246
Díaz AM, Alonso MLS, Gutiérrez MRV (2007) Biological traits of stream macroinvertebrates from a semi-arid catchment: patterns along complex environmental gradients. Freshw Biol 53(1):1–21
Ding N, Yang W, Zhou Y, Gonzalez-Bergonzoni I, Zhang J, Chen K, Vidal N, Jeppesen E, Liu Z, Wang B (2017) Different responses of functional traits and diversity of stream macroinvertebrates to environmental and spatial factors in the Xishuangbanna watershed of the upper Mekong River Basin, China. Sci Total Environ 574:288–299
Dolédec S, Chessel D, Ter Braak CJF, Champely S (1996) Matching species traits to environmental variables: a new three-table ordination method. Environ Ecol Stat 3:143–166
Edegbene AO, Elakhame LA, Arimoro FO, Osimen EC, Odume ON (2019) Development of macroinvertebrate multimetric index for ecological evaluation of a river in North Central Nigeria. Environ Monit Assess 191:274
Edegbene OA, Arimoro FO, Odume ON (2020) How does urban pollution influence macroinvertebrate traits in forested riverine systems? Water 12(11):3111. https://doi.org/10.3390/w12113111
Edegbene AO, Odume ON, Arimoro FO et al (2021) Identifying and classifying macroinvertebrate indicator signature traits and ecological preferences along urban pollution gradient in the Niger Delta. Environ Pollut 281:117076
Edegbene AO, Elakhame LA, Arimoro FO, Osimen EC, Akamagwuna FC, Edegbene Ovie TT et al (2023) Preliminary studies on macroinvertebrate biomonitoring of freshwater systems in the Afrotropics: a case study of the Chinchaga River in the Lower Niger-Benue Ecoregion of Nigeria. Biologia. https://doi.org/10.1007/s11756-023-01445-z
Fasinmirin JT, Olorunfemi IE (2014) Soil erodibility estimation under different tillage systems assessment in the rain forest climate of Osun State, Nigeria. Inte J Plant Soil Sci 3(1):16–35
Field CK, Fernandes MR, Ferreira MT, Hering D, Ormerod SJ, Venohr M, Gutiérrez-Cánovas C (2018) Evaluating riparian solutions to multiple stressor problems in river ecosystems—a conceptual study. Water Res 139:381–394
Forio MAE, Goethals PLM, Lock K, Asio V, Bande M, Thas O (2018) Model-based analysis of the relationship between macroinvertebrate traits and environmental river conditions. Environ Model Softw 106:57–67
Freeman LA, Corbett DR, Fitzgerald AM, Lemley DA, Quigg A, Steppe CN (2019) Impacts of urbanization and development on estuarine ecosystems and water quality. Estuaries Coasts 42:1821–1838
Friberg N, Bonada N, Bradley DC, Dunbar MJ, Edwards FK, Grey J, Hayes RB, Hildrew AG, Lamouroux N, Trimmer M et al (2010) Biomonitoring of human impacts in freshwater ecosystems: the good, the bad, and the ugly. Adv Ecol Res 43:1–68. https://doi.org/10.1016/S0065-2504(10)43001-3
Gordon ND, McMahon TA, Finlayson BL (1994) Stream hydrology, an introduction for ecologists. Wiley, New York
Jayawardana JMCK, Gunawardana WDTM, Udayakumara EPN, Westbrooke M (2017) Land use impacts on river health of Uma Oya, Sri Lanka: implications of spatial scales. Environ Monit Assess 189:192
Jones KK (2019) Investigating respiration and gas exchange in the aquatic bug Aphelocheirus aestivalis (Hemiptera: Aphelocheiridae), and surface dwelling and subterranean diving beetles (Coleoptera: Dytiscidae). Thesis, School of Biological Sciences: Ecology and Environmental Science, University of Adelaide
Keke UN, Arimoro FO, Ayanwale AV, Odume ON, Edegbene AO (2020) Biodiversity patterns along seasonality and environmental factors of stream macroinvertebrate communities of North-Central Nigeria. Egypt J Aquat Biol Fish 24(4):521–534. https://doi.org/10.21608/EJABF.2020.102102
Kennedy KA, Addison PA (1987) Some considerations for the use of visual estimates of plant cover in biomonitoring. J Ecol 75(1):151–157
Kilonzo F, Masese FO, Griensven AV, Bauwens W, Obando J, Lens PNL (2014) Spatial–temporal variability in water quality and macro-invertebrate assemblages in the Upper Mara River basin, Kenya. Phys Chem Earth Parts a/b/c 67–69:93–104
Kuzmanovic M, Doledec S, deCatro-Catala N, Ginebreda A, Sabater S, Munoz I, Barcelo D (2017) Environmental stressors as driver of the trait composition of benthic macroinvertebrates assemblages in polluted Iberian rivers. Environ Res 156:485–493
Lamouroux N, Dolédec S, Gayraud S (2004) Biological traits of stream macroinvertebrate communities: effects of microhabitat, reach, and basin filters. J N Am Benthol Soc 23:449–466
Lasisi M, Popoola A, Adediji A, Adedeji O, Babalola K (2017) City expansion and agricultural land loss within the peri-urban area of Osun State. Nigeria Ghana J Geogr 9(3):132–163
Leslie AW, Lamp WO (2017) Taxonomic and functional group composition of macroinvertebrate assemblages in agricultural drainage ditches. Hydrobiologia 787:99–110
Li F, Chung N, Bae MJ, Kwon YS, Park YS (2012) Relationships between stream macroinvertebrates and environmental variables at multiple spatial scales. Freshw Biol 57(10):2107–2124
Ligeiro R, Hughes RM, Kaufmann PR, Macedo DR, Firmiano KR, Ferreira WR, Oliveira D, Melo AS, Callisto M (2013) Defining quantitative stream disturbance gradients and the additive role of habitat variation to explain macroinvertebrate taxa richness. Ecol Ind 25:45–57
Malmqvist B, Rundle S (2002) Threats to the running water ecosystems of the world. Environ Conserv 29(2):134–153
Marques NCS, Jankowski KJ, Macedo MN, Juen L, Luiza-Andrade A, Deegan LA (2021) Riparian forests buffer the negative effects of cropland on macroinvertebrate diversity in lowland Amazonian stream. Hydrobiologia 848:3503–3520
Martínez HQ, Castro AR (2007) Aquatic insects as predators of mosquito larvae. J Am Mosq Control Assoc 23(sp2):110–117. https://doi.org/10.2987/8756-971X
Masese FO, Muchiri M, Raburu PO (2009) Macroinvertebrate assemblages as biological indicators of water quality in the Moiben River, Kenya. Afr J Aquat Sci 34(1):15–26
McGill BJ, Enquist BJ, Weiher E, Westoby M (2006) Rebuilding community ecology from functional traits. Trends Ecol Evol. https://doi.org/10.1016/j.tree.2006.02.002
Menezes S, Baird DJ, Soares AMVM (2010) Beyond taxonomy: a review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring. J Appl Ecol 47(4):711–719
Merritt RW, Cummins KW (1996) An introduction to the aquatic insects of North America. Kendall Hunt, St. Louis
Mohammed YM, Arimoro FO, Ayanwale AV, Adamu KM, Keke UN, Abubakar MD, Achebe AC (2021) The current state of water quality and benthic macroinvertebrate fauna in Chikke Stream (North Central Nigeria). Ukrainian J Ecol 11(3):26–34
De Moor IJ, Day JA, de Moor FC (2003) Guides to the freshwater invertebrates of southern Africa. Vol 7: Insecta I. Ephemeroptera, odonata and plecoptera. Water Research Commission, Report No. TT207/03
Murphy JF, Jones JI, Arnold A, Duerdoth CP, Pretty JL, Naden PS, Sear DA, Collins AL (2017) Can macroinvertebrate biological traits indicate fine-grained sediment conditions in streams? River Res Appl 33:1606–1617
Mwedzi T, Bere T, Mangadze T (2016) Macroinvertebrate assemblages in agricultural, mining, and urban tropical streams: implications for conservation and management. Environ Sci Pollut Res 23:11181–11192
Ngodhe SO, Raburu PO, Achieng A (2014) The impact of water quality on species diversity and richness of macroinvertebrates in small water bodies in Lake Victoria Basin, Kenya. J Ecol Nat Environ 6(1):32–41
Ochieng H, Okomu JO, Odong R (2019) Taxonomic challenges associated with identification guides of benthic macroinvertebrates for biomonitoring freshwater bodies in East Africa: a review. Afr J Aquat Sci 44(2):113–126
Odume ON (2020) Searching for urban pollution signature and sensitive macroinvertebrate traits and ecological preferences in a river in the Eastern Cape of South Africa. Ecol Ind 108:105759
Odume ON, Muller WJ, Arimoro FO, Palmer CG (2012) The impact of water quality deterioration on macroinvertebrate communities in the Swartkops River, South Africa: a multimetric approach. Afr J Aquat Sci 37(2):191–200
Odume ON, Palmer CG, Arimoro FO, Mensah PK (2014) Patterns of chironomid body-size distribution in an effluent impacted river in the Eastern Cape, South Africa. Afr J Aquat Sci 39(4):377–386
Odume ON, Palmer CG, Arimoro FO, Mensah PK (2016) Chironomid assemblage structure and morphological response to pollution in an effluent-impacted river, Eastern Cape, South Africa. Ecol Ind 67:391–402
Odume O, Ntloko P, Akamagwuna F, Dallas H, Barber-James H (2018) A trait database for South African macroinvertebrates. Unpublished WRC Report, Pretoria
Ogundari K, Ojo SO (2006) An examination of technical, economic and allocative efficiency of small farms: the case study of cassava farmers in Osun State of Nigeria. J Cent Eur Agric 7(3):423–432
Oksanen J, Blanchet D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Steven MHM, Szoecs E, Wagner H (2019) R. Development Core Team. Vegan: Community Ecology Package. https://cran.r-project.org. Accessed 11/02/2019. https://github.com/veganders/vegan
Omogbai BE (2010) Prediction of Northern Nigeria rainfall using sea surface temperature. J Hum Ecol 32(2):127–133
Onoharigho FO, Akpodimo EO, Edo GI (2022) The effect of uncontrolled dumping of solid waste on groundwater in Osun State, Nigeria. Fine Chem Eng 3:156–170
Padgham J, Jabbour J, Dietrich K (2015) Managing change and building resilience: a multi-stressor analysis of urban and peri-urban agriculture in Africa and Asia. Urban Clim 12:183–204
Pallottini M, Cappelletti D, Fabrizi A, Gaino E, Goretti E, Selvaggi R, Cereghino R (2017) Macroinvertebrate functional trait responses to chemical pollution in agricultural landscapes. River Res Appl 33:505–513
Pandey V, Jha DK, Kumar PS, Santhanakumar J, Venkatnarayanan S, Jebakumar JPP et al (2022) Effect of multiple stressors on the functional traits of sub-tidal macrobenthic fauna: a case study of the southeast coast of India. Mar Pollut Bull 175:113355. https://doi.org/10.1016/j.marpolbul.2022.113355
Poff NL, Olden JD, Vieira NKM, Finn DS, Simmons MP, Konddratieff BC (2006) Functional trait niches of North American lotic insects: traits-based ecological Applications in light of phylogenetic relationships. J N Am Benthol so 25:730–755
Qin G, Liu J, Xu S, Wang T (2020) Water quality assessment and pollution source apportionment in a highly regulated river of Northeast China. Environ Monit Assess 192:446
Reynolds J, Souty-Grosset C, Richardson A (2013) Ecological roles of crayfish in freshwater and terrestrial habitats. Freshwr Crayfish 19(2):197–218
Statzner B, Beche L (2010) Can biological invertebrate traits resolve effects of multiple stressors on running water ecosystems? Freshw Biol 55:80–199
Tomanova S, Usseglio-Polatera P (2007) Patterns of benthic community traits in neotropical streams: relationship to mesoscale spatial variability. Fundam Appl Limnol 170:243–255
Tomanova S, Moya N, Oberdorff T (2008) Using macroinvertebrate biological traits for assessing biotic integrity of neotropical streams. River Res Appl 24(9):1230–1239
Townsend CR, Hildrew AG (1994) Species traits in relation to a habitat templet for river systems. Freshw Biol 31(3):265–275
Vogel S (2008) Modes and scaling in aquatic locomotion. Integr Comp Biol 48(6):702–712
Wantzen KM, Mol JH (2013) Soil erosion from agriculture and mining: a threat to tropical stream ecosystems. Agriculture 3(4):660–683
Williams-Subiza EA, Brand C (2021) Functional response of benthic macroinvertebrates to fire disturbance in Patagonian streams. Hydrobiologia 848:1575–1591
Wurtsbaugh WA, Paerl HW, Dodds WK (2019) Nutrients, eutrophication and harmful algal blooms along the freshwater to marine continuum. WIREsWATER 6(5):e1373
Xu M, Wang Z, Duan X, Pan B (2014) Effects of pollution on macroinvertebrates and water quality bio-assessment. Hydrobiologia 729:247–259
Yetter SE (2012) Freshwater macroinvertebrates of the mid-atlantic region. mid-atlantic freshwater wetlands: advances in wetlands science, management, policy, and practice, pp 339–379
Cummins KW (1974) Structure and function of stream ecosystem. BioScience 24(11):631–641
Acknowledgements
We extend our gratitude to the staff members of Hydrobiology and Conservation Unit at the Department of Zoology, Obafemi Awolowo University, Ile-Ife, for generously providing access to their laboratory facilities and equipment for this study.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
Oluwaseun T. Akinpelu (conceptualization, methodology, monitoring, writing, reviewing and editing). Francis O. Arimoro (investigation, supervision and monitoring, formal analysis, reviewing and editing). Adesola V. Ayanwale (investigation, formal analysis, supervision). Victoria I. Chukwuemeka (investigation, formal analysis supervision). Tolulope A. Oladeji (methodology, formal analysis, draft preparation). Emmanuel O. Akindele (conceptualization, supervision and monitoring, reviewing and editing).
Corresponding author
Ethics declarations
Conflict of interest
The Authors declared that they have no competing interest.
Ethical approval
Not applicable.
Consent for publication
Not applicable.
Consent to participate
Not applicable.
Additional information
Handling editor: Maria J. Feio, PhD.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Akinpelu, O.T., Arimoro, F.O., Ayanwale, A.V. et al. Habitat characteristics and anthropogenic activities influence the distribution of macroinvertebrate traits and ecological preferences in Nigerian streams: a case study of Osun State, Nigeria. Aquat Ecol (2024). https://doi.org/10.1007/s10452-024-10108-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10452-024-10108-3